
In biology, a spore is a unit of sexual or asexual reproduction that may be adapted for dispersal and for survival, often for extended periods of time, in unfavourable conditions. Spores form part of the life cycles of many plants, algae, fungi and protozoa.

Zygomycota, or zygote fungi, is a former division or phylum of the kingdom Fungi. The members are now part of two phyla: the Mucoromycota and Zoopagomycota. Approximately 1060 species are known. They are mostly terrestrial in habitat, living in soil or on decaying plant or animal material. Some are parasites of plants, insects, and small animals, while others form symbiotic relationships with plants. Zygomycete hyphae may be coenocytic, forming septa only where gametes are formed or to wall off dead hyphae. Zygomycota is no longer recognised as it was not believed to be truly monophyletic.

An appressorium is a specialized cell typical of many fungal plant pathogens that is used to infect host plants. It is a flattened, hyphal "pressing" organ, from which a minute infection peg grows and enters the host, using turgor pressure capable of punching through even Mylar.

Basidiobolus ranarum is a filamentous fungus with worldwide distribution. The fungus was first isolated by Eidam in 1886. It can saprophytically live in the intestines of mainly cold-blooded vertebrates and on decaying fruits and soil. The fungus prefers glucose as a carbon source and grows rapidly at room temperature. Basidiobolus ranarum is also known as a cause of subcutaneous zygomycosis, usually causing granulomatous infections on a host's limbs. Infections are generally geographically limited to tropical and subtropical regions such as East and West Africa. Subcutaneous zygomycosis caused by B. ranarum is a rare disease and predominantly affects children and males. Common subcutaneous zygomycosis shows characteristic features and is relatively easy to be diagnosed; while, certain rare cases might show non-specific clinical features that might pose a difficulty on its identification. Although disease caused by this fungus is known to resolve spontaneously on its own, there are a number of treatments available.

Phytophthora palmivora is an oomycete that causes bud-rot of palms, fruit-rot or kole-roga of coconut and areca nut. These are among the most serious diseases caused by fungi and moulds in South India. It occurs almost every year in Malnad, Mysore, North & South Kanara, Malabar and other areas. Similar diseases of palms are also known to occur in Sri Lanka, Mauritius, and Sumatra. The causative organism was first identified as P. palmivora by Edwin John Butler in 1917.

The Entomophthorales are an order of fungi that were previously classified in the class Zygomycetes. A new subdivision, Entomophthoromycotina, in 2007, was circumscribed for them.
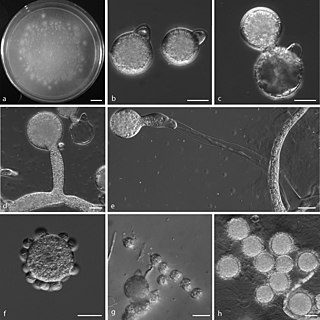
Conidiobolomycosis is a rare long-term fungal infection that is typically found just under the skin of the nose, sinuses, cheeks and upper lips. It may present with a nose bleed or a blocked or runny nose. Typically there is a firm painless swelling which can slowly extend to the nasal bridge and eyes, sometimes causing facial disfigurement.

Aspergillus terreus, also known as Aspergillus terrestris, is a fungus (mold) found worldwide in soil. Although thought to be strictly asexual until recently, A. terreus is now known to be capable of sexual reproduction. This saprotrophic fungus is prevalent in warmer climates such as tropical and subtropical regions. Aside from being located in soil, A. terreus has also been found in habitats such as decomposing vegetation and dust. A. terreus is commonly used in industry to produce important organic acids, such as itaconic acid and cis-aconitic acid, as well as enzymes, like xylanase. It was also the initial source for the drug mevinolin (lovastatin), a drug for lowering serum cholesterol.

Albugo is a genus of plant-parasitic oomycetes. Those are not true fungi (Eumycota), although many discussions of this organism still treat it as a fungus. The taxonomy of this genus is incomplete, but several species are plant pathogens. Albugo is one of three genera currently described in the family Albuginaceae, the taxonomy of many species is still in flux.

Colletotrichum lindemuthianum is a fungus which causes anthracnose, or black spot disease, of the common bean plant. It is considered a hemibiotrophic pathogen because it spends part of its infection cycle as a biotroph, living off of the host but not harming it, and the other part as a necrotroph, killing and obtaining nutrients from the host tissues.
Entomophthora is a fungal genus in the family Entomophthoraceae. Species in this genus are parasitic on flies and other two-winged insects. The genus was circumscribed by German physician Johann Baptist Georg Wolfgang Fresenius (1808–1866) in 1856.
Entomophthora muscae is a species of pathogenic fungus in the order Entomophthorales which causes a fatal disease in flies. It can cause epizootic outbreaks of disease in houseflies and has been investigated as a potential biological control agent.

Purpureocillium lilacinum is a species of filamentous fungus in the family Ophiocordycipitaceae. It has been isolated from a wide range of habitats, including cultivated and uncultivated soils, forests, grassland, deserts, estuarine sediments and sewage sludge, and insects. It has also been found in nematode eggs, and occasionally from females of root-knot and cyst nematodes. In addition, it has frequently been detected in the rhizosphere of many crops. The species can grow at a wide range of temperatures – from 8 to 38 °C for a few isolates, with optimal growth in the range 26 to 30 °C. It also has a wide pH tolerance and can grow on a variety of substrates. P. lilacinum has shown promising results for use as a biocontrol agent to control the growth of destructive root-knot nematodes.

Lichtheimia corymbifera is a thermophilic fungus in the phylum Zygomycota. It normally lives as a saprotrophic mold, but can also be an opportunistic pathogen known to cause pulmonary, CNS, rhinocerebral, or cutaneous infections in animals and humans with impaired immunity.

Actinoplanes italicus is distinguished by the cherry-red color of its vegetative mycelium, and by the production of soluble pigments. It is also known to produce sporangia when cultured on starch or skim milk agar. Very few strains have been found and cultured, thus A. italicus is relatively uncharacterized.

Rhizopus oryzae is a filamentous heterothallic microfungus that occurs as a saprotroph in soil, dung, and rotting vegetation. This species is very similar to Rhizopus stolonifer, but it can be distinguished by its smaller sporangia and air-dispersed sporangiospores. It differs from R. oligosporus and R. microsporus by its larger columellae and sporangiospores. The many strains of R. oryzae produce a wide range of enzymes such as carbohydrate digesting enzymes and polymers along with a number of organic acids, ethanol and esters giving it useful properties within the food industries, bio-diesel production, and pharmaceutical industries. It is also an opportunistic pathogen of humans causing mucormycosis.

Rhizopus stolonifer is commonly known as black bread mold. It is a member of Zygomycota and considered the most important species in the genus Rhizopus. It is one of the most common fungi in the world and has a global distribution although it is most commonly found in tropical and subtropical regions. It is a common agent of decomposition of stored foods. Like other members of the genus Rhizopus, R. stolonifer grows rapidly, mostly in indoor environments.

Mucor circinelloides is a dimorphic fungus belonging to the Order Mucorales. It has a worldwide distribution, found mostly in soil, dung and root vegetables. This species is described as not known to be able to produce mycotoxins, however it has been frequently reported to infect animals such as cattle and swine, as well as fowl, platypus and occasionally humans. Ketoacidotic patients are particularly at risk for infection by M. circinelloides.

Alternaria brassicicola is a fungal necrotrophic plant pathogen that causes black spot disease on a wide range of hosts, particularly in the genus of Brassica, including a number of economically important crops such as cabbage, Chinese cabbage, cauliflower, oilseeds, broccoli and canola. Although mainly known as a significant plant pathogen, it also contributes to various respiratory allergic conditions such as asthma and rhinoconjunctivitis. Despite the presence of mating genes, no sexual reproductive stage has been reported for this fungus. In terms of geography, it is most likely to be found in tropical and sub-tropical regions, but also in places with high rain and humidity such as Poland. It has also been found in Taiwan and Israel. Its main mode of propagation is vegetative. The resulting conidia reside in the soil, air and water. These spores are extremely resilient and can overwinter on crop debris and overwintering herbaceous plants.

Mortierella polycephala is a saprotrophic fungus with a wide geographical distribution occurring in many different habitats from soil and plants to salt marshes and slate slopes. It is the type species of the genus Mortierella, and was first described in 1863 by Henri Coemans. A characteristic feature of the fungus is the presence of stylospores, which are aerial, spiny resting spores (chlamydospores).
