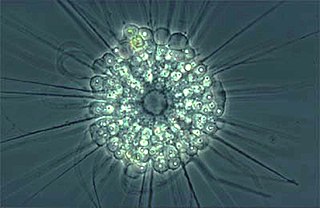
The actinophryids are an order of heliozoa, a polyphyletic array of stramenopiles, having a close relationship with pedinellids and Ciliophrys. They are common in fresh water and occasionally found in marine and soil habitats. Actinophryids are unicellular and roughly spherical in shape, with many axopodia that radiate outward from the cell body. Axopodia are a type of pseudopodia that are supported by hundreds of microtubules arranged in interlocking spirals and forming a needle-like internal structure or axoneme. Small granules, extrusomes, that lie under the membrane of the body and axopodia capture flagellates, ciliates and small metazoa that make contact with the arms.
Stramenopile is a clade of organisms distinguished by the presence of stiff tripartite external hairs. In most species, the hairs are attached to flagella, in some they are attached to other areas of the cellular surface, and in some they have been secondarily lost. Stramenopiles represent one of the three major clades in the SAR supergroup, along with Alveolata and Rhizaria.

Euglena is a genus of single cell flagellate eukaryotes. It is the best known and most widely studied member of the class Euglenoidea, a diverse group containing some 54 genera and at least 200 species. Species of Euglena are found in fresh water and salt water. They are often abundant in quiet inland waters where they may bloom in numbers sufficient to color the surface of ponds and ditches green (E. viridis) or red (E. sanguinea).
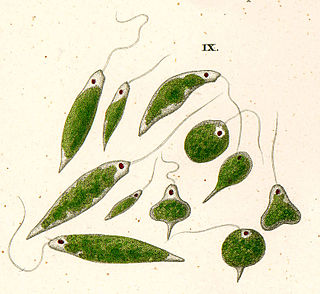
Euglenids are one of the best-known groups of flagellates, which are excavate eukaryotes of the phylum Euglenophyta and their cell structure is typical of that group. They are commonly found in freshwater, especially when it is rich in organic materials, with a few marine and endosymbiotic members. Many euglenids feed by phagocytosis, or strictly by diffusion. A monophyletic group consisting of the mixotrophic Rapaza viridis and the two groups Eutreptiales and Euglenales have chloroplasts and produce their own food through photosynthesis. This group is known to contain the carbohydrate paramylon.

Myxosporea is a class of microscopic parasites, belonging to the Myxozoa clade within Cnidaria. They have a complex life cycle which comprises vegetative forms in two hosts, an aquatic invertebrate and an ectothermic vertebrate, usually a fish. Each host releases a different type of spore. The two forms of spore are so different that until relatively recently they were treated as belonging to different classes within the Myxozoa.
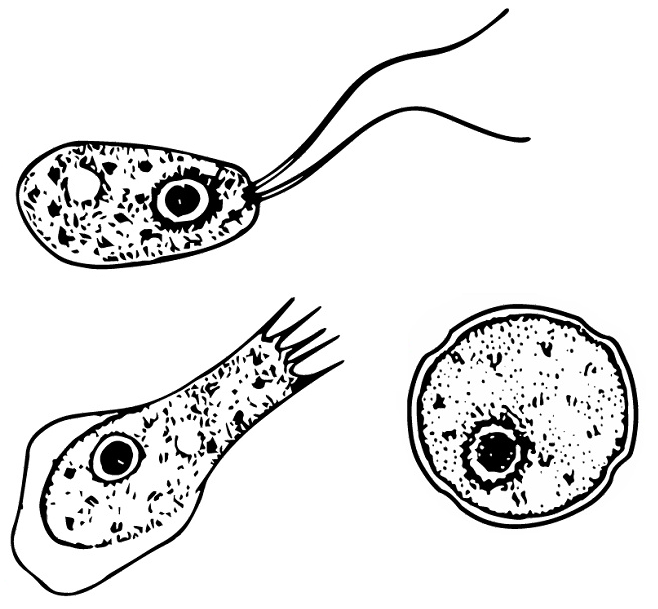
Naegleria is a free living amoebae protist genus consisting of 47 described species often found in warm aquatic environments as well as soil habitats worldwide. It has three life cycle forms: the amoeboid stage, the cyst stage, and the flagellated stage, and has been routinely studied for its ease in change from amoeboid to flagellated stages. The Naegleria genera became famous when Naegleria fowleri, a human pathogenic strain and the causative agent of primary amoebic meningoencephalitis (PAM), was discovered in 1965. Most species in the genus, however, are non pathogenic.

The Labyrinthulomycetes (ICBN) or Labyrinthulea (ICZN) are a class of protists that produce a network of filaments or tubes, which serve as tracks for the cells to glide along and absorb nutrients for them. The two main groups are the labyrinthulids and thraustochytrids. They are mostly marine, commonly found as parasites on algae and seagrasses or as decomposers on dead plant material. They also include some parasites of marine invertebrates.
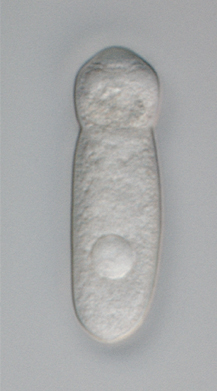
The gregarines are a group of Apicomplexan alveolates, classified as the Gregarinasina or Gregarinia. The large parasites inhabit the intestines of many invertebrates. They are not found in any vertebrates. However, gregarines are closely related to both Toxoplasma and Plasmodium, which cause toxoplasmosis and malaria, respectively. Both protists use protein complexes similar to those that are formed by the gregarines for gliding motility and invading target cells. This makes them excellent models for studying gliding motility with the goal of developing treatment options for toxoplasmosis and malaria. Thousands of different species of gregarines are expected to be found in insects, and 99% of these gregarines still need to be described. Each insect can be the host of multiple species. One of the most studied gregarines is Gregarina garnhami. In general, gregarines are regarded as very successful parasites, as their hosts are spread over the entire planet.
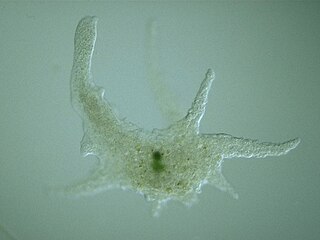
Chaos is a genus of single-celled amoeboid organisms in the family Amoebidae. The largest and best-known species, the so-called "giant amoeba" Chaos carolinensis, can reach lengths of 5 mm, although most specimens fall between 1 and 3 mm.
Members of the genus Selenomonas are referred to trivially as selenomonads. The genus Selenomonas constitutes a group of motile crescent-shaped bacteria and includes species living in the gastrointestinal tracts of animals, in particular the ruminants. A number of smaller forms discovered with the light microscope are now in culture but many, especially the large selenomonads are not, owing to their fastidious and incompletely known growth requirements.
In botany, a zoid or zoïd is a reproductive cell that possesses one or more flagella, and is capable of independent movement. Zoid can refer to either an asexually reproductive spore or a sexually reproductive gamete. In sexually reproductive gametes, zoids can be either male or female depending on the species. For example, some brown alga (Phaeophyceae) reproduce by producing multi-flagellated male and female gametes that recombine to form the diploid sporangia. Zoids are primarily found in some protists, diatoms, green alga, brown alga, non-vascular plants, and a few vascular plants. The most common classification group that produces zoids is the heterokonts or stramenopiles. These include green alga, brown alga, oomycetes, and some protists. The term is generally not used to describe motile, flagellated sperm found in animals. Zoid is also commonly confused for zooid which is a single organism that is part of a colonial animal.
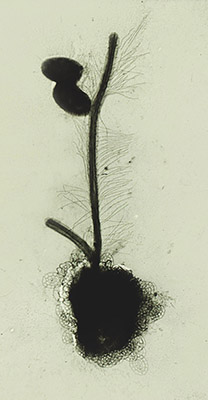
The genus Labyrinthula is part of the protist group Labyrinthulomycetes and contains thirteen species. The major feature of this genus is the formation of an ectoplasmic net secreted by specialized organelles called bothrosomes which surrounds the colony, which is also used by Labyrinthula for moving. The protist reproduces by zoosporulation as it sets some flagellated spores free from a sporangium. Zoospores prove the belonging of Labyrinthula in the Heterokont phylum due to the distinct flagellar morphology, in which the anterior one is covered in mastigonemes.

Peranema is a genus of free-living phagotrophic euglenids. There are more than 20 nominal species, varying in size between 8 and 200 micrometers. Peranema cells are gliding flagellates found in freshwater lakes, ponds and ditches, and are often abundant at the bottom of stagnant pools rich in decaying organic material. Although they belong to the class Euglenoidea, and are morphologically similar to the green Euglena, Peranema have no chloroplasts, and do not conduct autotrophy. Instead, they capture live prey, such as yeast, bacteria and other flagellates, consuming them with the help of a rigid feeding apparatus called a "rod-organ." Unlike the green euglenids, they lack both an eyespot (stigma), and the paraflagellar body (photoreceptor) that is normally coupled with that organelle. However, while Peranema lack a localized photoreceptor, they do possess the light-sensitive protein rhodopsin, and respond to changes in light with a characteristic "curling behaviour."
Parvilucifera is a genus of marine alveolates that parasitise dinoflagellates. Parvilucifera is a parasitic genus described in 1999 by Norén et al. It is classified perkinsozoa in the supraphylum of Alveolates. This taxon serves as a sister taxon to the dinoflagellates and apicomplexans. Thus far, five species have been described in this taxon, which include: P.infectans, P.sinerae, P.corolla, P.rostrata, and P.prorocentri. The genus Parvilucifera is morphologically characterized by flagellated zoospore. The life cycle of the species in this genus consist of free-living zoospores, an intracellular stage called trophont, and asexual division to form resting sporangium inside host cell. This taxon has gained more interest in research due to its potential significance in terms of negative regulation for dinoflagellates blooms, that have proved harmful for algal species, humans, and the shellfish industry.
The Eugregarinorida are the most large and diverse order of gregarines — parasitic protists belonging to the phylum Apicomplexa. Eugregarines are found in marine, freshwater and terrestrial habitats. These species possess large trophozoites that are significantly different in morphology and behavior from the sporozoites. This taxon contains most of the known gregarine species.
Schizochytrium is a genus of unicellular eukaryotes in the family Thraustochytriaceae, which are found in coastal marine habitats. They are assigned to the Stramenopiles (heterokonts), a group which also contains kelp and various microalgae.
Globisporangium sylvaticum is a plant pathogen, an oomycete known to cause root rot and damping off in a multitude of species. These species include apples, carrot, cherry laurel, cress, cucumber, garlic, lettuce, pea, rhododendron, and spinach. Symptoms of infection include stunting, wilt, chlorosis, and browning and eventual necrosis of roots. The pathogen can by identified by the presence of thick, microscopic, round spores within the cells of the root.
Nephromyces is a genus of apicomplexans that are symbionts of the ascidian genus Molgula.

Sphaeroforma arctica, is a unicellular eukaryote with a pivotal position in the tree of life. It was first isolated from the arctic marine amphipod Gammarus setosus. Like other Ichthyosporeans such as Creolimax and Abeoforma, Sphaeroforma arctica are spherical cells characterized with their capacity to grow into multi-nucleated coenocytes. However, a unique feature of S. arctica, is that no obvious budding, hyphal, amoeboid, sporal or flagellated growth stages have been observed in laboratory growth conditions.

Thraustochytrids are single-celled saprotrophic eukaryotes (decomposers) that are widely distributed in marine ecosystems, and which secrete enzymes including, but not limited to amylases, proteases, phosphatases. They are most abundant in regions with high amounts of detritus and decaying plant material. They play an important ecological role in mangroves, where they aid in nutrient cycling by decomposing decaying matter. Additionally, they contribute significantly to the synthesis of omega-3 polyunsaturated fatty acids (PUFAs): docosahexaenoic acid (DHA), and eicosapentaenoic acid (EPA), which are essential fatty acids for the growth and reproduction of crustaceans. Thraustochytrids are members of the class Labyrinthulea, a group of protists that had previously been incorrectly categorized as fungi due to their similar appearance and lifestyle. With the advent of DNA sequencing technology, labyrinthulomycetes were appropriately placed with other stramenopiles and subsequently categorized as a group of Labyrinthulomycetes.

