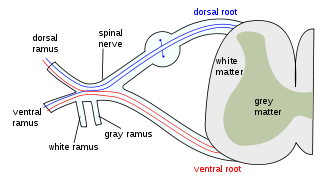
Grey matter, or gray matter in American English, is a major component of the central nervous system, consisting of neuronal cell bodies, neuropil, glial cells, synapses, and capillaries. Grey matter is distinguished from white matter in that it contains numerous cell bodies and relatively few myelinated axons, while white matter contains relatively few cell bodies and is composed chiefly of long-range myelinated axons. The colour difference arises mainly from the whiteness of myelin. In living tissue, grey matter actually has a very light grey colour with yellowish or pinkish hues, which come from capillary blood vessels and neuronal cell bodies.

Image registration is the process of transforming different sets of data into one coordinate system. Data may be multiple photographs, data from different sensors, times, depths, or viewpoints. It is used in computer vision, medical imaging, military automatic target recognition, and compiling and analyzing images and data from satellites. Registration is necessary in order to be able to compare or integrate the data obtained from these different measurements.

Functional magnetic resonance imaging or functional MRI (fMRI) measures brain activity by detecting changes associated with blood flow. This technique relies on the fact that cerebral blood flow and neuronal activation are coupled. When an area of the brain is in use, blood flow to that region also increases.
Neuroscience and intelligence refers to the various neurological factors that are partly responsible for the variation of intelligence within species or between different species. A large amount of research in this area has been focused on the neural basis of human intelligence. Historic approaches to studying the neuroscience of intelligence consisted of correlating external head parameters, for example head circumference, to intelligence. Post-mortem measures of brain weight and brain volume have also been used. More recent methodologies focus on examining correlates of intelligence within the living brain using techniques such as magnetic resonance imaging (MRI), functional MRI (fMRI), electroencephalography (EEG), positron emission tomography and other non-invasive measures of brain structure and activity.

Morphometrics or morphometry refers to the quantitative analysis of form, a concept that encompasses size and shape. Morphometric analyses are commonly performed on organisms, and are useful in analyzing their fossil record, the impact of mutations on shape, developmental changes in form, covariances between ecological factors and shape, as well for estimating quantitative-genetic parameters of shape. Morphometrics can be used to quantify a trait of evolutionary significance, and by detecting changes in the shape, deduce something of their ontogeny, function or evolutionary relationships. A major objective of morphometrics is to statistically test hypotheses about the factors that affect shape.
Functional integration is the study of how brain regions work together to process information and effect responses. Though functional integration frequently relies on anatomic knowledge of the connections between brain areas, the emphasis is on how large clusters of neurons – numbering in the thousands or millions – fire together under various stimuli. The large datasets required for such a whole-scale picture of brain function have motivated the development of several novel and general methods for the statistical analysis of interdependence, such as dynamic causal modelling and statistical linear parametric mapping. These datasets are typically gathered in human subjects by non-invasive methods such as EEG/MEG, fMRI, or PET. The results can be of clinical value by helping to identify the regions responsible for psychiatric disorders, as well as to assess how different activities or lifestyles affect the functioning of the brain.

Analysis of Functional NeuroImages (AFNI) is an open-source environment for processing and displaying functional MRI data—a technique for mapping human brain activity.
In neuroimaging, spatial normalization is an image processing step, more specifically an image registration method. Human brains differ in size and shape, and one goal of spatial normalization is to deform human brain scans so one location in one subject's brain scan corresponds to the same location in another subject's brain scan.

Talairach coordinates, also known as Talairach space, is a 3-dimensional coordinate system of the human brain, which is used to map the location of brain structures independent from individual differences in the size and overall shape of the brain. It is still common to use Talairach coordinates in functional brain imaging studies and to target transcranial stimulation of brain regions. However, alternative methods such as the MNI Coordinate System have largely replaced Talairach for stereotaxy and other procedures.

Voxel-based morphometry is a computational approach to neuroanatomy that measures differences in local concentrations of brain tissue, through a voxel-wise comparison of multiple brain images.
Statistical shape analysis is an analysis of the geometrical properties of some given set of shapes by statistical methods. For instance, it could be used to quantify differences between male and female gorilla skull shapes, normal and pathological bone shapes, leaf outlines with and without herbivory by insects, etc. Important aspects of shape analysis are to obtain a measure of distance between shapes, to estimate mean shapes from samples, to estimate shape variability within samples, to perform clustering and to test for differences between shapes. One of the main methods used is principal component analysis (PCA). Statistical shape analysis has applications in various fields, including medical imaging, computer vision, computational anatomy, sensor measurement, and geographical profiling.
Gyrification is the process of forming the characteristic folds of the cerebral cortex.
Seed-based d mapping or SDM is a statistical technique created by Joaquim Radua for meta-analyzing studies on differences in brain activity or structure which used neuroimaging techniques such as fMRI, VBM, DTI or PET. It may also refer to a specific piece of software created by the SDM Project to carry out such meta-analyses.
Karl John Friston FRS FMedSci FRSB is a British neuroscientist and theoretician at University College London. He is an authority on brain imaging and theoretical neuroscience, especially the use of physics-inspired statistical methods to model neuroimaging data and other random dynamical systems. Friston is a key architect of the free energy principle and active inference. In imaging neuroscience he is best known for statistical parametric mapping and dynamic causal modelling. In October 2022, he joined VERSES Inc, a California-based cognitive computing company focusing on artificial intelligence designed using the principles of active inference, as Chief Scientist.
Medical image computing (MIC) is an interdisciplinary field at the intersection of computer science, information engineering, electrical engineering, physics, mathematics and medicine. This field develops computational and mathematical methods for solving problems pertaining to medical images and their use for biomedical research and clinical care.
The parieto-frontal integration theory (P-FIT) considers intelligence to relate to how well different brain regions integrate to form intelligent behaviors. The theory proposes that large scale brain networks connect brain regions, including regions within frontal, parietal, temporal, and cingulate cortices, underlie the biological basis of human intelligence. These regions, which overlap significantly with the task-positive network, allow the brain to communicate and exchange information efficiently with one another. Support for this theory is primarily based on neuroimaging evidence, with support from lesion studies. The P-FIT is influential in that it explains the majority of current neuroimaging findings, as well as increasing empirical support for cognition being the result of large-scale brain networks, rather than numerous domain-specific processes or modules. A 2010 review of the neuroscience of intelligence described P-FIT as "the best available answer to the question of where in the brain intelligence resides".
Computational anatomy is an interdisciplinary field of biology focused on quantitative investigation and modelling of anatomical shapes variability. It involves the development and application of mathematical, statistical and data-analytical methods for modelling and simulation of biological structures.
Large deformation diffeomorphic metric mapping (LDDMM) is a specific suite of algorithms used for diffeomorphic mapping and manipulating dense imagery based on diffeomorphic metric mapping within the academic discipline of computational anatomy, to be distinguished from its precursor based on diffeomorphic mapping. The distinction between the two is that diffeomorphic metric maps satisfy the property that the length associated to their flow away from the identity induces a metric on the group of diffeomorphisms, which in turn induces a metric on the orbit of shapes and forms within the field of Computational Anatomy. The study of shapes and forms with the metric of diffeomorphic metric mapping is called diffeomorphometry.
Joaquim Radua is a Spanish psychiatrist and developer of methods for meta-analysis of neuroimaging studies. He has been named as one of the most cited researchers in Psychiatry / Psychology.
Arterial spin labeling (ASL), also known as arterial spin tagging, is a magnetic resonance imaging technique used to quantify cerebral blood perfusion by labelling blood water as it flows throughout the brain. ASL specifically refers to magnetic labeling of arterial blood below or in the imaging slab, without the need of gadolinium contrast. A number of ASL schemes are possible, the simplest being flow alternating inversion recovery (FAIR) which requires two acquisitions of identical parameters with the exception of the out-of-slice saturation; the difference in the two images is theoretically only from inflowing spins, and may be considered a 'perfusion map'. The ASL technique was developed by Alan P. Koretsky, Donald S. Williams, John A. Detre and John S. Leigh Jr in 1992.
