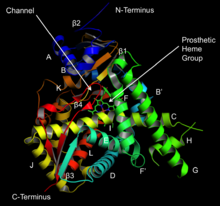
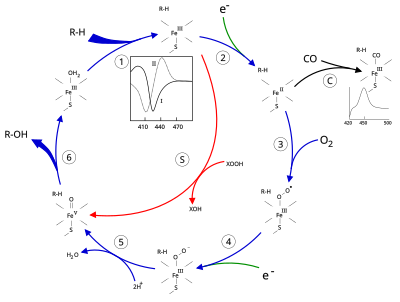
Cytochrome P450 2E1 is a member of the cytochrome P450 mixed-function oxidase system, which is involved in the metabolism of xenobiotics in the body. This class of enzymes is divided up into a number of subcategories, including CYP1, CYP2, and CYP3, which as a group are largely responsible for the breakdown of foreign compounds in mammals.
Drug metabolism is the metabolic breakdown of drugs by living organisms, usually through specialized enzymatic systems. More generally, xenobiotic metabolism is the set of metabolic pathways that modify the chemical structure of xenobiotics, which are compounds foreign to an organism's normal biochemistry, such as any drug or poison. These pathways are a form of biotransformation present in all major groups of organisms and are considered to be of ancient origin. These reactions often act to detoxify poisonous compounds. The study of drug metabolism is called pharmacokinetics.

In biochemistry, flavin adenine dinucleotide (FAD) is a redox-active coenzyme associated with various proteins, which is involved with several enzymatic reactions in metabolism. A flavoprotein is a protein that contains a flavin group, which may be in the form of FAD or flavin mononucleotide (FMN). Many flavoproteins are known: components of the succinate dehydrogenase complex, α-ketoglutarate dehydrogenase, and a component of the pyruvate dehydrogenase complex.

Thromboxane A synthase 1 , also known as TBXAS1, is a cytochrome P450 enzyme that, in humans, is encoded by the TBXAS1 gene.
Any enzyme system that includes cytochrome P450 protein or domain can be called a P450-containing system.

Cytochrome P450 17A1 is an enzyme of the hydroxylase type that in humans is encoded by the CYP17A1 gene on chromosome 10. It is ubiquitously expressed in many tissues and cell types, including the zona reticularis and zona fasciculata of the adrenal cortex as well as gonadal tissues. It has both 17α-hydroxylase and 17,20-lyase activities, and is a key enzyme in the steroidogenic pathway that produces progestins, mineralocorticoids, glucocorticoids, androgens, and estrogens. More specifically, the enzyme acts upon pregnenolone and progesterone to add a hydroxyl (-OH) group at carbon 17 position (C17) of the steroid D ring, or acts upon 17α-hydroxyprogesterone and 17α-hydroxypregnenolone to split the side-chain off the steroid nucleus.

Cytochrome P450 reductase is a membrane-bound enzyme required for electron transfer from NADPH to cytochrome P450 and other heme proteins including heme oxygenase in the endoplasmic reticulum of the eukaryotic cell.

In enzymology, a camphor 5-monooxygenase (EC 1.14.15.1) is an enzyme that catalyzes the chemical reaction

Cholesterol 24-hydroxylase, also commonly known as cholesterol 24S-hydroxylase, cholesterol 24-monooxygenase, CYP46, or CYP46A1, is an enzyme that catalyzes the conversion of cholesterol to 24S-hydroxycholesterol. It is responsible for the majority of cholesterol turnover in the human central nervous system. The systematic name of this enzyme class is cholesterol,NADPH:oxygen oxidoreductase (24-hydroxylating).
In enzymology, an unspecific monooxygenase (EC 1.14.14.1) is an enzyme that catalyzes the chemical reaction
Cytochrome P450 3A5 is a protein that in humans is encoded by the CYP3A5 gene.

Cytochrome P450 4F2 is a protein that in humans is encoded by the CYP4F2 gene. This protein is an enzyme, a type of protein that catalyzes chemical reactions inside cells. This specific enzyme is part of the superfamily of cytochrome P450 (CYP) enzymes, and the encoding gene is part of a cluster of cytochrome P450 genes located on chromosome 19.

CYP20A1 is a protein which in humans is encoded by the CYP20A1 gene.

(+)-Menthofuran synthase (EC 1.14.13.104, menthofuran synthase, (+)-pulegone 9-hydroxylase, (+)-MFS, cytochrome P450 menthofuran synthase) is an enzyme with systematic name (+)-pulegone,NADPH:oxygen oxidoreductase (9-hydroxylating). This enzyme catalyses the following chemical reaction
Tryptophan N-monooxygenase (EC 1.14.13.125, tryptophan N-hydroxylase, CYP79B1, CYP79B2, CYP79B3) is an enzyme with systematic name L-tryptophan,NADPH:oxygen oxidoreductase (N-hydroxylating). This enzyme catalyses the following chemical reaction
1,8-Cineole 2-endo-monooxygenase (EC 1.14.14.133, Formerly EC 1.14.13.156, P450cin, CYP176A, CYP176A1) is an enzyme with systematic name 1,8-cineole,NADPH:oxygen oxidoreductase (2-endo-hydroxylating). This enzyme catalyses the following chemical reaction
Cytochrome P450 omega hydroxylases, also termed cytochrome P450 ω-hydroxylases, CYP450 omega hydroxylases, CYP450 ω-hydroxylases, CYP omega hydroxylase, CYP ω-hydroxylases, fatty acid omega hydroxylases, cytochrome P450 monooxygenases, and fatty acid monooxygenases, are a set of cytochrome P450-containing enzymes that catalyze the addition of a hydroxyl residue to a fatty acid substrate. The CYP omega hydroxylases are often referred to as monoxygenases; however, the monooxygenases are CYP450 enzymes that add a hydroxyl group to a wide range of xenobiotic and naturally occurring endobiotic substrates, most of which are not fatty acids. The CYP450 omega hydroxylases are accordingly better viewed as a subset of monooxygenases that have the ability to hydroxylate fatty acids. While once regarded as functioning mainly in the catabolism of dietary fatty acids, the omega oxygenases are now considered critical in the production or break-down of fatty acid-derived mediators which are made by cells and act within their cells of origin as autocrine signaling agents or on nearby cells as paracrine signaling agents to regulate various functions such as blood pressure control and inflammation.
This article covers protein engineering of cytochrome (CYP) P450 enzymes. P450s are involved in a range of biochemical catabolic and anabolic process. Natural P450s can perform several different types of chemical reactions including hydroxylations, N,O,S-dealkylations, epoxidations, sulfoxidations, aryl-aryl couplings, ring contractions and expansions, oxidative cyclizations, alcohol/aldehyde oxidations, desaturations, nitrogen oxidations, decarboxylations, nitrations, as well as oxidative and reductive dehalogenations. Engineering efforts often strive for 1) improved stability 2) improved activity 3) improved substrate scope 4) enabled ability to catalyze unnatural reactions. P450 engineering is an emerging field in the areas of chemical biology and synthetic organic chemistry (chemoenzymatic).
Cytochrome P450 aromatic O-demethylase is a bacterial enzyme that catalyzes the demethylation of lignin and various lignols. The net reaction follows the following stoichiometry, illustrated with a generic methoxy arene:
In biochemistry, cytochrome P450 enzymes have been identified in all kingdoms of life: animals, plants, fungi, protists, bacteria, and archaea, as well as in viruses. As of 2018, more than 300,000 distinct CYP proteins are known.