Gene knockouts are a widely used genetic engineering technique that involves the targeted removal or inactivation of a specific gene within an organism's genome. This can be done through a variety of methods, including homologous recombination, CRISPR-Cas9, and TALENs.
Gene knockdown is an experimental technique by which the expression of one or more of an organism's genes is reduced. The reduction can occur either through genetic modification or by treatment with a reagent such as a short DNA or RNA oligonucleotide that has a sequence complementary to either gene or an mRNA transcript.

CRISPR is a family of DNA sequences found in the genomes of prokaryotic organisms such as bacteria and archaea. These sequences are derived from DNA fragments of bacteriophages that had previously infected the prokaryote. They are used to detect and destroy DNA from similar bacteriophages during subsequent infections. Hence these sequences play a key role in the antiviral defense system of prokaryotes and provide a form of acquired immunity. CRISPR is found in approximately 50% of sequenced bacterial genomes and nearly 90% of sequenced archaea.

RNA editing is a molecular process through which some cells can make discrete changes to specific nucleotide sequences within an RNA molecule after it has been generated by RNA polymerase. It occurs in all living organisms and is one of the most evolutionarily conserved properties of RNAs. RNA editing may include the insertion, deletion, and base substitution of nucleotides within the RNA molecule. RNA editing is relatively rare, with common forms of RNA processing not usually considered as editing. It can affect the activity, localization as well as stability of RNAs, and has been linked with human diseases.

A kinetoplast is a network of circular DNA inside a mitochondrion that contains many copies of the mitochondrial genome. The most common kinetoplast structure is a disk, but they have been observed in other arrangements. Kinetoplasts are only found in Excavata of the class Kinetoplastida. The variation in the structures of kinetoplasts may reflect phylogenic relationships between kinetoplastids. A kinetoplast is usually adjacent to the organism's flagellar basal body, suggesting that it is bound to some components of the cytoskeleton. In Trypanosoma brucei this cytoskeletal connection is called the tripartite attachment complex and includes the protein p166.

Genome editing, or genome engineering, or gene editing, is a type of genetic engineering in which DNA is inserted, deleted, modified or replaced in the genome of a living organism. Unlike early genetic engineering techniques that randomly inserts genetic material into a host genome, genome editing targets the insertions to site-specific locations. The basic mechanism involved in genetic manipulations through programmable nucleases is the recognition of target genomic loci and binding of effector DNA-binding domain (DBD), double-strand breaks (DSBs) in target DNA by the restriction endonucleases, and the repair of DSBs through homology-directed recombination (HDR) or non-homologous end joining (NHEJ).
In molecular biology, trans-activating CRISPR RNA (tracrRNA) is a small trans-encoded RNA. It was first discovered by Emmanuelle Charpentier in her study of the human pathogen Streptococcus pyogenes, a type of bacteria that causes harm to humanity. In bacteria and archaea, CRISPR-Cas constitute an RNA-mediated defense system that protects against viruses and plasmids. This defensive pathway has three steps. First, a copy of the invading nucleic acid is integrated into the CRISPR locus. Next, CRISPR RNAs (crRNAs) are transcribed from this CRISPR locus. The crRNAs are then incorporated into effector complexes, where the crRNA guides the complex to the invading nucleic acid and the Cas proteins degrade this nucleic acid. There are several CRISPR system subtypes.

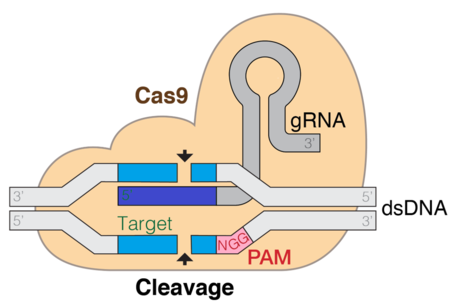
Cas9 is a 160 kilodalton protein which plays a vital role in the immunological defense of certain bacteria against DNA viruses and plasmids, and is heavily utilized in genetic engineering applications. Its main function is to cut DNA and thereby alter a cell's genome. The CRISPR-Cas9 genome editing technique was a significant contributor to the Nobel Prize in Chemistry in 2020 being awarded to Emmanuelle Charpentier and Jennifer Doudna.

CRISPR interference (CRISPRi) is a genetic perturbation technique that allows for sequence-specific repression of gene expression in prokaryotic and eukaryotic cells. It was first developed by Stanley Qi and colleagues in the laboratories of Wendell Lim, Adam Arkin, Jonathan Weissman, and Jennifer Doudna. Sequence-specific activation of gene expression refers to CRISPR activation (CRISPRa).
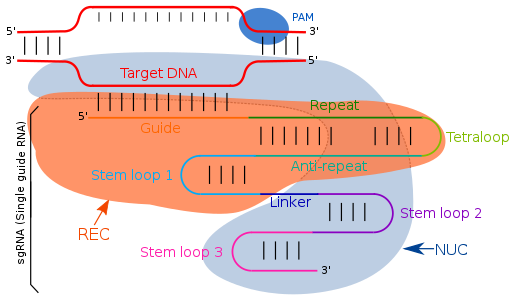
A protospacer adjacent motif (PAM) is a 2–6-base pair DNA sequence immediately following the DNA sequence targeted by the Cas9 nuclease in the CRISPR bacterial adaptive immune system. The PAM is a component of the invading virus or plasmid, but is not found in the bacterial host genome and hence is not a component of the bacterial CRISPR locus. Cas9 will not successfully bind to or cleave the target DNA sequence if it is not followed by the PAM sequence. PAM is an essential targeting component which distinguishes bacterial self from non-self DNA, thereby preventing the CRISPR locus from being targeted and destroyed by the CRISPR-associated nuclease.
CRISPR-Cas design tools are computer software platforms and bioinformatics tools used to facilitate the design of guide RNAs (gRNAs) for use with the CRISPR/Cas gene editing system.

Cas12a is a subtype of Cas12 proteins and an RNA-guided endonuclease that forms part of the CRISPR system in some bacteria and archaea. It originates as part of a bacterial immune mechanism, where it serves to destroy the genetic material of viruses and thus protect the cell and colony from viral infection. Cas12a and other CRISPR associated endonucleases use an RNA to target nucleic acid in a specific and programmable matter. In the organisms from which it originates, this guide RNA is a copy of a piece of foreign nucleic acid that previously infected the cell.
A cryptogene is a gene that has had its transcript edited. This phenomenon is observed in various organisms, particularly in Kinetoplastids and Myxomycetes, where the process plays a crucial role in mitochondrial gene expression.
No-SCAR genome editing is an editing method that is able to manipulate the Escherichia coli genome. The system relies on recombineering whereby DNA sequences are combined and manipulated through homologous recombination. No-SCAR is able to manipulate the E. coli genome without the use of the chromosomal markers detailed in previous recombineering methods. Instead, the λ-Red recombination system facilitates donor DNA integration while Cas9 cleaves double-stranded DNA to counter-select against wild-type cells. Although λ-Red and Cas9 genome editing are widely used technologies, the no-SCAR method is novel in combining the two functions; this technique is able to establish point mutations, gene deletions, and short sequence insertions in several genomic loci with increased efficiency and time sensitivity.
CRISPR activation (CRISPRa) is a type of CRISPR tool that uses modified versions of CRISPR effectors without endonuclease activity, with added transcriptional activators on dCas9 or the guide RNAs (gRNAs).
Off-target genome editing refers to nonspecific and unintended genetic modifications that can arise through the use of engineered nuclease technologies such as: clustered, regularly interspaced, short palindromic repeats (CRISPR)-Cas9, transcription activator-like effector nucleases (TALEN), meganucleases, and zinc finger nucleases (ZFN). These tools use different mechanisms to bind a predetermined sequence of DNA (“target”), which they cleave, creating a double-stranded chromosomal break (DSB) that summons the cell's DNA repair mechanisms and leads to site-specific modifications. If these complexes do not bind at the target, often a result of homologous sequences and/or mismatch tolerance, they will cleave off-target DSB and cause non-specific genetic modifications. Specifically, off-target effects consist of unintended point mutations, deletions, insertions inversions, and translocations.

CRISPR gene editing standing for "Clustered Regularly Interspaced Short Palindromic Repeats" is a genetic engineering technique in molecular biology by which the genomes of living organisms may be modified. It is based on a simplified version of the bacterial CRISPR-Cas9 antiviral defense system. By delivering the Cas9 nuclease complexed with a synthetic guide RNA (gRNA) into a cell, the cell's genome can be cut at a desired location, allowing existing genes to be removed and/or new ones added in vivo.
Prime editing is a 'search-and-replace' genome editing technology in molecular biology by which the genome of living organisms may be modified. The technology directly writes new genetic information into a targeted DNA site. It uses a fusion protein, consisting of a catalytically impaired Cas9 endonuclease fused to an engineered reverse transcriptase enzyme, and a prime editing guide RNA (pegRNA), capable of identifying the target site and providing the new genetic information to replace the target DNA nucleotides. It mediates targeted insertions, deletions, and base-to-base conversions without the need for double strand breaks (DSBs) or donor DNA templates.
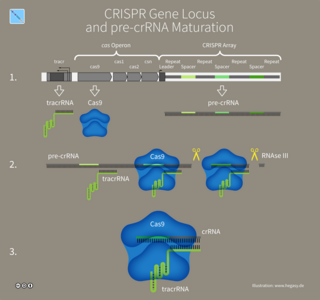
CRISPR RNA or crRNA is a RNA transcript from the CRISPR locus. CRISPR-Cas is an adaptive immune system found in bacteria and archaea to protect against mobile genetic elements, like viruses, plasmids, and transposons. The CRISPR locus contains a series of repeats interspaced with unique spacers. These unique spacers can be acquired from MGEs.
The Fanzor (Fz) protein is an eukaryotic, RNA-guided DNA endonuclease, which means it is a type of DNA cutting enzyme that uses RNA to target genes of interest. It has been recently discovered and explored in a number of studies. In bacteria, RNA-guided DNA endonuclease systems, such as the CRISPR/Cas system, serve as an immune system to prevent infection by cutting viral genetic material. Currently, CRISPR/Cas9-mediated's DNA cleavage has extensive application in biological research, and wide-reaching medical potential in human gene editing.