Chemokines, or chemotactic cytokines, are a family of small cytokines or signaling proteins secreted by cells that induce directional movement of leukocytes, as well as other cell types, including endothelial and epithelial cells. In addition to playing a major role in the activation of host immune responses, chemokines are important for biological processes, including morphogenesis and wound healing, as well as in the pathogenesis of diseases like cancers.
Cell adhesion molecules (CAMs) are a subset of cell surface proteins that are involved in the binding of cells with other cells or with the extracellular matrix (ECM), in a process called cell adhesion. In essence, CAMs help cells stick to each other and to their surroundings. CAMs are crucial components in maintaining tissue structure and function. In fully developed animals, these molecules play an integral role in generating force and movement and consequently ensuring that organs are able to execute their functions normally. In addition to serving as "molecular glue", CAMs play important roles in the cellular mechanisms of growth, contact inhibition, and apoptosis. Aberrant expression of CAMs may result in a wide range of pathologies, ranging from frostbite to cancer.

Selectin P ligand, also known as SELPLG or CD162, is a human gene.

The selectins are a family of cell adhesion molecules. All selectins are single-chain transmembrane glycoproteins that share similar properties to C-type lectins due to a related amino terminus and calcium-dependent binding. Selectins bind to sugar moieties and so are considered to be a type of lectin, cell adhesion proteins that bind sugar polymers.
P-selectin is a type-1 transmembrane protein that in humans is encoded by the SELP gene.

ICAM-1 also known as CD54 is a protein that in humans is encoded by the ICAM1 gene. This gene encodes a cell surface glycoprotein which is typically expressed on endothelial cells and cells of the immune system. It binds to integrins of type CD11a / CD18, or CD11b / CD18 and is also exploited by rhinovirus as a receptor for entry into respiratory epithelium.

E-selectin, also known as CD62 antigen-like family member E (CD62E), endothelial-leukocyte adhesion molecule 1 (ELAM-1), or leukocyte-endothelial cell adhesion molecule 2 (LECAM2), is a selectin cell adhesion molecule expressed only on endothelial cells activated by cytokines. Like other selectins, it plays an important part in inflammation. In humans, E-selectin is encoded by the SELE gene.

CD11c, also known as Integrin, alpha X (ITGAX), is a gene that encodes for CD11c.
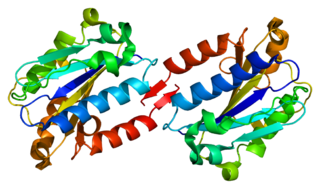
Integrin, alpha L , also known as ITGAL, is a protein that in humans is encoded by the ITGAL gene. CD11a functions in the immune system. It is involved in cellular adhesion and costimulatory signaling. It is the target of the drug efalizumab.
High endothelial venules (HEV) are specialized post-capillary venules characterized by plump endothelial cells as opposed to the usual flatter endothelial cells found in regular venules. HEVs enable lymphocytes circulating in the blood to directly enter a lymph node.
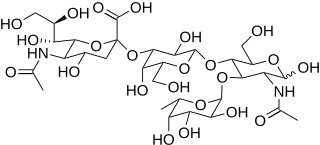
Sialyl LewisX (sLeX), also known as cluster of differentiation 15s (CD15s) or stage-specific embryonic antigen 1 (SSEA-1), is a tetrasaccharide carbohydrate which is usually attached to O-glycans on the surface of cells. It is known to play a vital role in cell-to-cell recognition processes. It is also the means by which an egg attracts sperm; first, to stick to it, then bond with it and eventually form a zygote.

Mucosal vascular addressin cell adhesion molecule 1 (MAdCAM-1) is a protein that in humans is encoded by the MADCAM1 gene. The protein encoded by this gene is an endothelial cell adhesion molecule that interacts preferentially with the leukocyte beta7 integrin LPAM-1, L-selectin, and VLA-4 on myeloid cells to direct leukocytes into mucosal and inflamed tissues. It is a member of the immunoglobulin superfamily and is similar to ICAM-1 and VCAM-1.

Chemokine ligand 21 (CCL21) is a small cytokine belonging to the CC chemokine family. This chemokine is also known as 6Ckine, exodus-2, and secondary lymphoid-tissue chemokine (SLC). CCL21 elicits its effects by binding to a cell surface chemokine receptor known as CCR7. The main function of CCL21 is to guide CCR7 expressing leukocytes to the secondary lymphoid organs, such as lymph nodes and Peyer´s patches.
Glycosylation-dependent cell adhesion molecule-1 (GLYCAM1) is a proteoglycan ligand expressed on cells of the high endothelial venules in lymphoid tissues. It is the ligand for the receptor L-selectin allowing for naive lymphocytes to exit the bloodstream into lymphoid tissues. GLYCAM1 binds to L-selectin by presenting one or more O-linked carbohydrates to the lectin domain of the leukocyte cell surface selectin. Data suggests that GLYCAM1 is a hormone-regulated milk protein that is part of the milk mucin complex.
Lymphocyte homing receptors are cell adhesion molecules expressed on lymphocyte cell membranes that recognize addressins on target tissues. Lymphocyte homing refers to adhesion of the circulating lymphocytes in blood to specialized endothelial cells within lymphoid organs. These diverse tissue-specific adhesion molecules on lymphocytes and on endothelial cells contribute to the development of specialized immune responses.

Leukocyte extravasation is the movement of leukocytes out of the circulatory system and towards the site of tissue damage or infection. This process forms part of the innate immune response, involving the recruitment of non-specific leukocytes. Monocytes also use this process in the absence of infection or tissue damage during their development into macrophages.
Alpha-(1,3)-fucosyltransferase is an enzyme that in humans is encoded by the FUT7 gene.
Lymph node stromal cells are essential to the structure and function of the lymph node whose functions include: creating an internal tissue scaffold for the support of hematopoietic cells; the release of small molecule chemical messengers that facilitate interactions between hematopoietic cells; the facilitation of the migration of hematopoietic cells; the presentation of antigens to immune cells at the initiation of the adaptive immune system; and the homeostasis of lymphocyte numbers. Stromal cells originate from multipotent mesenchymal stem cells.
Gut-specific homing is the mechanism by which activated T cells and antibody-secreting cells (ASCs) are targeted to both inflamed and non-inflamed regions of the gut in order to provide an effective immune response. This process relies on the key interaction between the integrin α4β7 and the addressin MadCAM-1 on the surfaces of the appropriate cells. Additionally, this interaction is strengthened by the presence of CCR9, a chemokine receptor, which interacts with TECK. Vitamin A-derived retinoic acid regulates the expression of these cell surface proteins.
Integrin α4β7 is an integrin heterodimer composed of CD49d (alpha-4) subunit and beta-7 subunit noncovalently linked. LPAM-1 is expressed on the cell surface of leukocytes. This receptor is involved in lymphocyte trafficking pathway to site of inflammation in intestinal tissues.