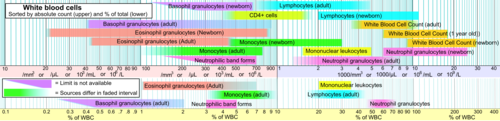
A cytotoxic T cell (also known as TC, cytotoxic T lymphocyte, CTL, T-killer cell, cytolytic T cell, CD8+ T-cell or killer T cell) is a T lymphocyte (a type of white blood cell) that kills cancer cells, cells that are infected by intracellular pathogens (such as viruses or bacteria), or cells that are damaged in other ways.

The T helper cells (Th cells), also known as CD4+ cells or CD4-positive cells, are a type of T cell that play an important role in the adaptive immune system. They aid the activity of other immune cells by releasing cytokines. They are considered essential in B cell antibody class switching, breaking cross-tolerance in dendritic cells, in the activation and growth of cytotoxic T cells, and in maximizing bactericidal activity of phagocytes such as macrophages and neutrophils. CD4+ cells are mature Th cells that express the surface protein CD4. Genetic variation in regulatory elements expressed by CD4+ cells determines susceptibility to a broad class of autoimmune diseases.

The adaptive immune system, also known as the acquired immune system, or specific immune system is a subsystem of the immune system that is composed of specialized, systemic cells and processes that eliminate pathogens or prevent their growth. The acquired immune system is one of the two main immunity strategies found in vertebrates.

The T-cell receptor (TCR) is a protein complex found on the surface of T cells, or T lymphocytes, that is responsible for recognizing fragments of antigen as peptides bound to major histocompatibility complex (MHC) molecules. The binding between TCR and antigen peptides is of relatively low affinity and is degenerate: that is, many TCRs recognize the same antigen peptide and many antigen peptides are recognized by the same TCR.
CD8 is a transmembrane glycoprotein that serves as a co-receptor for the T-cell receptor (TCR). Along with the TCR, the CD8 co-receptor plays a role in T cell signaling and aiding with cytotoxic T cell-antigen interactions.
A co-receptor is a cell surface receptor that binds a signalling molecule in addition to a primary receptor in order to facilitate ligand recognition and initiate biological processes, such as entry of a pathogen into a host cell.
Lck is a 56 kDa protein that is found inside specialized cells of the immune system called lymphocytes. The Lck is a member of Src kinase family (SFK) and is important for the activation of T-cell receptor (TCR) signaling in both naive T cells and effector T cells. The role of Lck is less prominent in the activation or in the maintenance of memory CD8 T cells in comparison to CD4 T cells. In addition, the constitutive activity of the mouse Lck homolog varies among memory T cell subsets. It seems that in mice, in the effector memory T cell (TEM) population, more than 50% of Lck is present in a constitutively active conformation, whereas less than 20% of Lck is present as active form in central memory T cells. These differences are due to differential regulation by SH2 domain–containing phosphatase-1 (Shp-1) and C-terminal Src kinase.

CD28 is one of the proteins expressed on T cells that provide co-stimulatory signals required for T cell activation and survival. T cell stimulation through CD28 in addition to the T-cell receptor (TCR) can provide a potent signal for the production of various interleukins.

In immunology, an immunological synapse is the interface between an antigen-presenting cell or target cell and a lymphocyte such as a T cell, B cell, or natural killer cell. The interface was originally named after the neuronal synapse, with which it shares the main structural pattern. An immunological synapse consists of molecules involved in T cell activation, which compose typical patterns—activation clusters. Immunological synapses are the subject of much ongoing research.

HLA class II histocompatibility antigen, DR alpha chain is a protein that in humans is encoded by the HLA-DRA gene. HLA-DRA encodes the alpha subunit of HLA-DR. Unlike the alpha chains of other Human MHC class II molecules, the alpha subunit is practically invariable. However it can pair with, in any individual, the beta chain from 3 different DR beta loci, DRB1, and two of any DRB3, DRB4, or DRB5 alleles. Thus there is the potential that any given individual can form 4 different HLA-DR isoforms.
MHC-restricted antigen recognition, or MHC restriction, refers to the fact that a T cell can interact with a self-major histocompatibility complex molecule and a foreign peptide bound to it, but will only respond to the antigen when it is bound to a particular MHC molecule.
In immunology, a naive T cell (Th0 cell) is a T cell that has differentiated in the thymus, and successfully undergone the positive and negative processes of central selection in the thymus. Among these are the naive forms of helper T cells (CD4+) and cytotoxic T cells (CD8+). Any naive T cell is considered immature and, unlike activated or memory T cells, has not encountered its cognate antigen within the periphery. After this encounter, the naive T cell is considered a mature T cell.

ZAP-70 is a protein normally expressed near the surface membrane of lymphocytes. It is most prominently known to be recruited upon antigen binding to the T cell receptor (TCR), and it plays a critical role in T cell signaling.

T-cell surface glycoprotein CD3 gamma chain is a protein that in humans is encoded by the CD3G gene.
CD83 is a human protein encoded by the CD83 gene.

Lymphocyte-activation gene 3, also known as LAG-3, is a protein which in humans is encoded by the LAG3 gene. LAG3, which was discovered in 1990 and was designated CD223 after the Seventh Human Leucocyte Differentiation Antigen Workshop in 2000, is a cell surface molecule with diverse biological effects on T cell function but overall has an immune inhibitory effect. It is an immune checkpoint receptor and as such is the target of various drug development programs by pharmaceutical companies seeking to develop new treatments for cancer and autoimmune disorders. In soluble form it is also being developed as a cancer drug in its own right.

CD8a, is a human gene.
Immunoevasins are proteins expressed by some viruses that enable the virus to evade immune recognition by interfering with MHC I complexes in the infected cell, therefore blocking the recognition of viral protein fragments by CD8+ cytotoxic T lymphocytes. Less frequently, MHC II antigen presentation and induced-self molecules may also be targeted. Some viral immunoevasins block peptide entry into the endoplasmic reticulum (ER) by targeting the TAP transporters. Immunoevasins are particularly abundant in viruses that are capable of establishing long-term infections of the host, such as herpesviruses.
Kinetic-segregation is a model proposed for the mechanism of T-cell receptor (TCR) triggering. It offers an explanation for how TCR binding to its ligand triggers T-cell activation, based on size-sensitivity for the molecules involved. Simon J. Davis and Anton van der Merwe, University of Oxford, proposed this model in 1996. According to the model, TCR signalling is initiated by segregation of phosphatases with large extracellular domains from the TCR complex when binding to its ligand, allowing small kinases to phosphorylate intracellular domains of the TCR without inhibition. Its might also be applicable to other receptors of the Non-catalytic tyrosine-phosphorylated receptors family such as CD28.

KIR2DL3, Killer cell immunoglobulin-like receptor 2DL3 is a transmembrane glycoprotein expressed by the natural killer cells and the subsets of the T cells. The KIR genes are polymorphic, which means that they have many different alleles. The KIR genes are also extremely homologous, which means that they are similar in position, structure and evolutionary origin, but not necessarily in function.