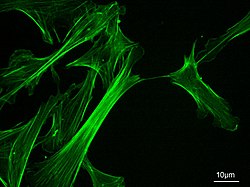
Microfilaments, also called actin filaments, are protein filaments in the cytoplasm of eukaryoticcells that form part of the cytoskeleton. They are primarily composed of polymers of actin, but are modified by and interact with numerous other proteins in the cell. Microfilaments are usually about 7 nm in diameter and made up of two strands of actin. Microfilament functions include cytokinesis, amoeboid movement, cell motility, changes in cell shape, endocytosis and exocytosis, cell contractility, and mechanical stability. Microfilaments are flexible and relatively strong, resisting buckling by multi-piconewton compressive forces and filament fracture by nanonewton tensile forces. In inducing cell motility, one end of the actin filament elongates while the other end contracts, presumably by myosin II molecular motors.[1] Additionally, they function as part of actomyosin-driven contractile molecular motors, wherein the thin filaments serve as tensile platforms for myosin's ATP-dependent pulling action in muscle contraction and pseudopod advancement. Microfilaments have a tough, flexible framework which helps the cell in movement.[2]
Actin was first discovered in rabbit skeletal muscle in the mid 1940 by F.B. Straub.[3] Almost 20 years later, H.E. Huxley demonstrated that actin is essential for muscle constriction. The mechanism in which actin creates long filaments was first described in the mid 1980. Later studies showed that actin has an important role in cell shape, motility, and cytokinesis.
Organization
Actin filaments are assembled in two general types of structures: bundles and networks. Bundles can be composed of polar filament arrays, in which all barbed ends point to the same end of the bundle, or non-polar arrays, where the barbed ends point towards both ends. A class of actin-binding proteins, called cross-linking proteins, dictate the formation of these structures. Cross-linking proteins determine filament orientation and spacing in the bundles and networks. These structures are regulated by many other classes of actin-binding proteins, including motor proteins, branching proteins, severing proteins, polymerization promoters, and capping proteins.
In vitro self-assembly
Measuring approximately 6 nm in diameter,[4] microfilaments are the thinnest fibers of the cytoskeleton. They are polymers of actin subunits (globular actin, or G-actin), which as part of the fiber are referred to as filamentous actin, or F-actin. Each microfilament is made up of two helical, interlaced strands of subunits. Much like microtubules, actin filaments are polarized. Electron micrographs have provided evidence of their fast-growing barbed-ends and their slow-growing pointed-end. This polarity has been determined by the pattern created by the binding of myosin S1 fragments: they themselves are subunits of the larger myosin II protein complex. The pointed end is commonly referred to as the minus (−) end and the barbed end is referred to as the plus (+) end.
In vitro actin polymerization, or nucleation, starts with the self-association of three G-actin monomers to form a trimer. ATP-bound actin then itself binds the barbed end, and the ATP is subsequently hydrolyzed. ATP hydrolysis occurs with a half time of about 2 seconds,[5] while the half time for the dissociation of the inorganic phosphate is about 6 minutes.[5] This autocatalyzed event reduces the binding strength between neighboring subunits, and thus generally destabilizes the filament. In vivo actin polymerization is catalyzed by a class of filament end-tracking molecular motors known as actoclampins. Recent evidence suggests that the rate of ATP hydrolysis and the rate of monomer incorporation are strongly coupled.
Subsequently, ADP-actin dissociates slowly from the pointed end, a process significantly accelerated by the actin-binding protein, cofilin. ADP bound cofilin severs ADP-rich regions nearest the (−)-ends. Upon release, the free actin monomer slowly dissociates from ADP, which in turn rapidly binds to the free ATP diffusing in the cytosol, thereby forming the ATP-actin monomeric units needed for further barbed-end filament elongation. This rapid turnover is important for the cell's movement. End-capping proteins such as CapZ prevent the addition or loss of monomers at the filament end where actin turnover is unfavorable, such as in the muscle apparatus.
Actin polymerization together with capping proteins were recently used to control the 3-dimensional growth of protein filament so as to perform 3D topologies useful in technology and the making of electrical interconnect. Electrical conductivity is obtained by metallisation of the protein 3D structure.[6][7]
Mechanism of force generation
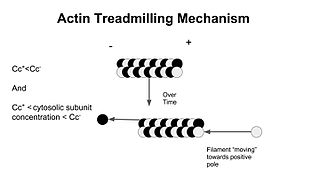
As a result of ATP hydrolysis, filaments elongate approximately 10 times faster at their barbed ends than their pointed ends. At steady-state, the polymerization rate at the barbed end matches the depolymerization rate at the pointed end, and microfilaments are said to be treadmilling. Treadmilling results in elongation in the barbed end and shortening in the pointed-end, so that the filament in total moves. Since both processes are energetically favorable, this means force is generated, the energy ultimately coming from ATP.[1]
Actin in cells
Intracellular actin cytoskeletal assembly and disassembly are tightly regulated by cell signaling mechanisms. Many signal transduction systems use the actin cytoskeleton as a scaffold, holding them at or near the inner face of the peripheral membrane. This subcellular location allows immediate responsiveness to transmembrane receptor action and the resulting cascade of signal-processing enzymes.
Because actin monomers must be recycled to sustain high rates of actin-based motility during chemotaxis, cell signalling is believed to activate cofilin, the actin-filament depolymerizing protein which binds to ADP-rich actin subunits nearest the filament's pointed-end and promotes filament fragmentation, with concomitant depolymerization in order to liberate actin monomers. In most animal cells, monomeric actin is bound to profilin and thymosin beta-4, both of which preferentially bind with one-to-one stoichiometry to ATP-containing monomers. Although thymosin beta-4 is strictly a monomer-sequestering protein, the behavior of profilin is far more complex. Profilin enhances the ability of monomers to assemble by stimulating the exchange of actin-bound ADP for solution-phase ATP to yield actin-ATP and ADP. Profilin is transferred to the leading edge by virtue of its PIP2 binding site, and it employs its poly-L-proline binding site to dock onto end-tracking proteins. Once bound, profilin-actin-ATP is loaded into the monomer-insertion site of actoclampin motors.[citation needed]
Another important component in filament formation is the Arp2/3 complex, which binds to the side of an already existing filament (or "mother filament"), where it nucleates the formation of a new daughter filament at a 70 degree angle relative to the mother filament, effecting a fan-like branched filament network.[8]
Specialized unique actin cytoskeletal structures are found adjacent to the plasma membrane. Four remarkable examples include red blood cells, human embryonic kidney cells, neurons, and sperm cells. In red blood cells, a spectrin-actin hexagonal lattice is formed by interconnected short actin filaments.[9] In human embryonic kidney cells, the cortical actin forms a scale-free fractal structure.[10] First found in neuronal axons, actin forms periodic rings that are stabilized by spectrin and adducin[11][12]– and this ring structure was then found by He et al 2016 to occur in almost every neuronal type and glial cells, across seemingly every animal taxon including Caenorhabditis elegans, Drosophila, Gallus gallus and Mus musculus.[13] And in mammalian sperm, actin forms a helical structure in the midpiece, i.e., the first segment of the flagellum.[14]
Associated proteins
In non-muscle cells, actin filaments are formed proximal to membrane surfaces. Their formation and turnover are regulated by many proteins, including:
Actin depolymerizing proteins such as ADF/cofilin.
The actin filament network in non-muscle cells is highly dynamic. The actin filament network is arranged with the barbed-end of each filament attached to the cell's peripheral membrane by means of clamped-filament elongation motors, the above-mentioned "actoclampins", formed from a filament barbed-end and a clamping protein (formins, VASP, Mena, WASP, and N-WASP).[15] The primary substrate for these elongation motors is profilin-actin-ATP complex which is directly transferred to elongating filament ends.[16] The pointed-end of each filament is oriented toward the cell's interior. In the case of lamellipodial growth, the Arp2/3 complex generates a branched network, and in filopodia a parallel array of filaments is formed.
Actin acts as a track for myosin motor motility
Myosin motors are intracellular ATP-dependent enzymes that bind to and move along actin filaments. Various classes of myosin motors have very different behaviors, including exerting tension in the cell and transporting cargo vesicles.
A proposed model – actoclampins track filament ends
The term actoclampin is derived from acto- to indicate the involvement of an actin filament, as in actomyosin, and clamp to indicate a clasping device used for strengthening flexible/moving objects and for securely fastening two or more components, followed by the suffix -in to indicate its protein origin. An actin filament end-tracking protein may thus be termed a clampin.
Dickinson and Purich recognized that prompt ATP hydrolysis could explain the forces achieved during actin-based motility.[15] They proposed a simple mechanoenzymatic sequence known as the Lock, Load & Fire Model, in which an end-tracking protein remains tightly bound ("locked" or clamped) onto the end of one sub-filament of the double-stranded actin filament. After binding to Glycyl-Prolyl-Prolyl-Prolyl-Prolyl-Prolyl-registers on tracker proteins, Profilin-ATP-actin is delivered ("loaded") to the unclamped end of the other sub-filament, whereupon ATP within the already clamped terminal subunit of the other subfragment is hydrolyzed ("fired"), providing the energy needed to release that arm of the end-tracker, which then can bind another Profilin-ATP-actin to begin a new monomer-addition round.[citation needed]
Steps involved
The following steps describe one force-generating cycle of an actoclampin molecular motor:
The polymerization cofactor profilin and the ATP·actin combine to form a profilin-ATP-actin complex that then binds to the end-tracking unit
The cofactor and monomer are transferred to the barbed-end of an actin already clamped filament
The tracking unit and cofactor dissociate from the adjacent protofilament, in a step that can be facilitated by ATP hydrolysis energy to modulate the affinity of the cofactor and/or the tracking unit for the filament; and this mechanoenzymatic cycle is then repeated, starting this time on the other sub-filament growth site.[citation needed]
When operating with the benefit of ATP hydrolysis, AC motors generate per-filament forces of 8–9 pN, which is far greater than the per-filament limit of 1–2 pN for motors operating without ATP hydrolysis.[15][17][18] The term actoclampin is generic and applies to all actin filament end-tracking molecular motors, irrespective of whether they are driven actively by an ATP-activated mechanism or passively.
Some actoclampins (e.g., those involving Ena/VASP proteins, WASP, and N-WASP) apparently require Arp2/3-mediated filament initiation to form the actin polymerization nucleus that is then "loaded" onto the end-tracker before processive motility can commence. To generate a new filament, Arp2/3 requires a "mother" filament, monomeric ATP-actin, and an activating domain from Listeria ActA or the VCA region of N-WASP. The Arp2/3 complex binds to the side of the mother filament, forming a Y-shaped branch having a 70 degree angle with respect to the longitudinal axis of the mother filament. Then upon activation by ActA or VCA, the Arp complex is believed to undergo a major conformational change, bringing its two actin-related protein subunits near enough to each other to generate a new filament gate. Whether ATP hydrolysis may be required for nucleation and/or Y-branch release is a matter under active investigation.
Related Research Articles
A myofibril is a basic rod-like organelle of a muscle cell. Skeletal muscles are composed of long, tubular cells known as muscle fibers, and these cells contain many chains of myofibrils. Each myofibril has a diameter of 1–2 micrometres. They are created during embryonic development in a process known as myogenesis.
Actin is a family of globular multi-functional proteins that form microfilaments in the cytoskeleton, and the thin filaments in muscle fibrils. It is found in essentially all eukaryotic cells, where it may be present at a concentration of over 100 μM; its mass is roughly 42 kDa, with a diameter of 4 to 7 nm.
Myosins are a superfamily of motor proteins best known for their roles in muscle contraction and in a wide range of other motility processes in eukaryotes. They are ATP-dependent and responsible for actin-based motility.
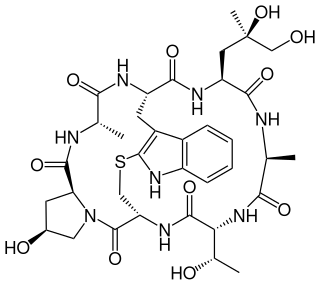
Phalloidin belongs to a class of toxins called phallotoxins, which are found in the death cap mushroom (Amanita phalloides). It is a rigid bicyclic heptapeptide that is lethal after a few days when injected into the bloodstream. The major symptom of phalloidin poisoning is acute hunger due to the destruction of liver cells. It functions by binding and stabilizing filamentous actin (F-actin) and effectively prevents the depolymerization of actin fibers. Due to its tight and selective binding to F-actin, derivatives of phalloidin containing fluorescent tags are used widely in microscopy to visualize F-actin in biomedical research.
Motor proteins are a class of molecular motors that can move along the cytoplasm of cells. They convert chemical energy into mechanical work by the hydrolysis of ATP. Flagellar rotation, however, is powered by a proton pump.
Profilin is an actin-binding protein involved in the dynamic turnover and reconstruction of the actin cytoskeleton. It is found in most eukaryotic organisms. Profilin is important for spatially and temporally controlled growth of actin microfilaments, which is an essential process in cellular locomotion and cell shape changes. This restructuring of the actin cytoskeleton is essential for processes such as organ development, wound healing, and the hunting down of infectious intruders by cells of the immune system.
The lamellipodium is a cytoskeletal protein actin projection on the leading edge of the cell. It contains a quasi-two-dimensional actin mesh; the whole structure propels the cell across a substrate. Within the lamellipodia are ribs of actin called microspikes, which, when they spread beyond the lamellipodium frontier, are called filopodia. The lamellipodium is born of actin nucleation in the plasma membrane of the cell and is the primary area of actin incorporation or microfilament formation of the cell.
ADF/cofilin is a family of actin-binding proteins associated with the rapid depolymerization of actin microfilaments that give actin its characteristic dynamic instability. This dynamic instability is central to actin's role in muscle contraction, cell motility and transcription regulation.
Cytochalasins are fungal metabolites that have the ability to bind to actin filaments and block polymerization and the elongation of actin. As a result of the inhibition of actin polymerization, cytochalasins can change cellular morphology, inhibit cellular processes such as cell division, and even cause cells to undergo apoptosis. Cytochalasins have the ability to permeate cell membranes, prevent cellular translocation and cause cells to enucleate. Cytochalasins can also have an effect on other aspects of biological processes unrelated to actin polymerization. For example, cytochalasin A and cytochalasin B can also inhibit the transport of monosaccharides across the cell membrane, cytochalasin H has been found to regulate plant growth, cytochalasin D inhibits protein synthesis and cytochalasin E prevents angiogenesis.
In molecular biology, treadmilling is a phenomenon observed within protein filaments of the cytoskeletons of many cells, especially in actin filaments and microtubules. It occurs when one end of a filament grows in length while the other end shrinks, resulting in a section of filament seemingly "moving" across a stratum or the cytosol. This is due to the constant removal of the protein subunits from these filaments at one end of the filament, while protein subunits are constantly added at the other end. Treadmilling was discovered by Wegner, who defined the thermodynamic and kinetic constraints. Wegner recognized that: “The equilibrium constant (K) for association of a monomer with a polymer is the same at both ends, since the addition of a monomer to each end leads to the same polymer.”; a simple reversible polymer can’t treadmill; ATP hydrolysis is required. GTP is hydrolyzed for microtubule treadmilling.
In biology, a protein filament is a long chain of protein monomers, such as those found in hair, muscle, or in flagella. Protein filaments form together to make the cytoskeleton of the cell. They are often bundled together to provide support, strength, and rigidity to the cell. When the filaments are packed up together, they are able to form three different cellular parts. The three major classes of protein filaments that make up the cytoskeleton include: actin filaments, microtubules and intermediate filaments.
CapZ, also known as CAPZ, CAZ1 and CAPPA1, is a capping protein that caps the barbed end of actin filaments in muscle cells.
Protein cordon-bleu is a protein that in humans is encoded by the COBL gene.
The Actin assembly-inducing protein (ActA) is a protein encoded and used by Listeria monocytogenes to propel itself through a mammalian host cell. ActA is a bacterial surface protein comprising a membrane-spanning region. In a mammalian cell the bacterial ActA interacts with the Arp2/3 complex and actin monomers to induce actin polymerization on the bacterial surface generating an actin comet tail. The gene encoding ActA is named actA or prtB.
Actin remodeling is the biochemical process that allows for the dynamic alterations of cellular organization. The remodeling of actin filaments occurs in a cyclic pattern on cell surfaces and exists as a fundamental aspect to cellular life. During the remodeling process, actin monomers polymerize in response to signaling cascades that stem from environmental cues. The cell's signaling pathways cause actin to affect intracellular organization of the cytoskeleton and often consequently, the cell membrane. Again triggered by environmental conditions, actin filaments break back down into monomers and the cycle is completed. Actin-binding proteins (ABPs) aid in the transformation of actin filaments throughout the actin remodeling process. These proteins account for the diverse structure and changes in shape of Eukaryotic cells. Despite its complexity, actin remodeling may result in complete cytoskeletal reorganization in under a minute.
mDia1 is a member of the protein family called the formins and is a Rho effector. It is the mouse version of the diaphanous homolog 1 of Drosophila. mDia1 localizes to cells' mitotic spindle and midbody, plays a role in stress fiber and filopodia formation, phagocytosis, activation of serum response factor, formation of adherens junctions, and it can act as a transcription factor. mDia1 accelerates actin nucleation and elongation by interacting with barbed ends of actin filaments. The gene encoding mDia1 is located on Chromosome 18 of Mus musculus and named Diap1.
Arp2/3 complex is a seven-subunit protein complex that plays a major role in the regulation of the actin cytoskeleton. It is a major component of the actin cytoskeleton and is found in most actin cytoskeleton-containing eukaryotic cells. Two of its subunits, the Actin-Related Proteins ARP2 and ARP3, closely resemble the structure of monomeric actin and serve as nucleation sites for new actin filaments. The complex binds to the sides of existing ("mother") filaments and initiates growth of a new ("daughter") filament at a distinctive 70 degree angle from the mother. Branched actin networks are created as a result of this nucleation of new filaments. The regulation of rearrangements of the actin cytoskeleton is important for processes like cell locomotion, phagocytosis, and intracellular motility of lipid vesicles.
In molecular biology, the cyclase-associated protein family (CAP) is a family of highly conserved actin-binding proteins present in a wide range of organisms including yeast, flies, plants, and mammals. CAPs are multifunctional proteins that contain several structural domains. CAP is involved in species-specific signalling pathways. In Drosophila, CAP functions in Hedgehog-mediated eye development and in establishing oocyte polarity. In Dictyostelium discoideum, CAP is involved in microfilament reorganisation near the plasma membrane in a PIP2-regulated manner and is required to perpetuate the cAMP relay signal to organise fruitbody formation. In plants, CAP is involved in plant signalling pathways required for co-ordinated organ expansion. In yeast, CAP is involved in adenylate cyclase activation, as well as in vesicle trafficking and endocytosis. In both yeast and mammals, CAPs appear to be involved in recycling G-actin monomers from ADF/cofilins for subsequent rounds of filament assembly. In mammals, there are two different CAPs that share 64% amino acid identity.
An actin nucleation core is a protein trimer with three actin monomers. It is called a nucleation core because it leads to the energetically favorable elongation reaction once a tetramer is formed from a trimer. Actin protein dimers and trimers are energetically unfavorable. Actin nucleators like the Arp2/3 complex of proteins from the formin family are most frequently involved in this process. Actin nucleation factors start the polymerization of actin within cells.
Edwin W. Taylor is an adjunct professor of cell and developmental biology at Northwestern University. He was elected to the National Academy of Sciences in 2001. Taylor received a BA in physics and chemistry from the University of Toronto in 1952; an MSc in physical chemistry from McMaster University in 1955, and a PhD in biophysics from the University of Chicago in 1957. In 2001 Taylor was elected to the National Academy of Scineces in Cellular and Developmental Biology and Biochemistry.
References
1 2 Roberts K, Raff M, Alberts B, Walter P, Lewis J, Johnson A (March 2002). Molecular Biology of the Cell (4thed.). Routledge. p.1616. ISBN0-8153-3218-1.
↑ Galland R, Leduc P, Guérin C, Peyrade D, Blanchoin L, Théry M (May 2013). "Fabrication of three-dimensional electrical connections by means of directed actin self-organization". Nature Materials. 12 (5): 416–21. Bibcode:2013NatMa..12..416G. doi:10.1038/nmat3569. PMID23396247.
↑ US Patent US 9070702, Method for obtaining three-dimensional actin structures and uses thereof, Jean-Christophe Gabriel, Laurent Blanchoin, Manuel Thery, Remi Galland
↑ Dickinson RB, Southwick FS, Purich DL (October 2002). "A direct-transfer polymerization model explains how the multiple profilin-binding sites in the actoclampin motor promote rapid actin-based motility". Archives of Biochemistry and Biophysics. 406 (2): 296–301. doi:10.1016/s0003-9861(02)00212-6. PMID12361718.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.