
A mycorrhiza is a symbiotic association between a fungus and a plant. The term mycorrhiza refers to the role of the fungus in the plant's rhizosphere, its root system. Mycorrhizae play important roles in plant nutrition, soil biology, and soil chemistry.

A truffle is the fruiting body of a subterranean ascomycete fungus, predominantly one of the many species of the genus Tuber. In addition to Tuber, over one hundred other genera of fungi are classified as truffles including Geopora, Peziza, Choiromyces, and Leucangium. These genera belong to the class Pezizomycetes and the Pezizales order. Several truffle-like basidiomycetes are excluded from Pezizales, including Rhizopogon and Glomus. Truffles are ectomycorrhizal fungi, so they are usually found in close association with tree roots. Spore dispersal is accomplished through fungivores, animals that eat fungi. These fungi have significant ecological roles in nutrient cycling and drought tolerance.

An endophyte is an endosymbiont, often a bacterium or fungus, that lives within a plant for at least part of its life cycle without causing apparent disease. Endophytes are ubiquitous and have been found in all species of plants studied to date; however, most of the endophyte/plant relationships are not well understood. Some endophytes may enhance host growth and nutrient acquisition and improve the plant's ability to tolerate abiotic stresses, such as drought, and decrease biotic stresses by enhancing plant resistance to insects, pathogens and herbivores. Although endophytic bacteria and fungi are frequently studied, endophytic archaea are increasingly being considered for their role in plant growth promotion as part of the core microbiome of a plant.

An arbuscular mycorrhiza (AM) is a type of mycorrhiza in which the symbiont fungus penetrates the cortical cells of the roots of a vascular plant forming arbuscules. Arbuscular mycorrhiza is a type of endomycorrhiza along with ericoid mycorrhiza and orchid mycorrhiza. They are characterized by the formation of unique tree-like structures, the arbuscules. In addition, globular storage structures called vesicles are often encountered.

Forest ecology is the scientific study of the interrelated patterns, processes, flora, fauna and ecosystems in forests. The management of forests is known as forestry, silviculture, and forest management. A forest ecosystem is a natural woodland unit consisting of all plants, animals, and micro-organisms in that area functioning together with all of the non-living physical (abiotic) factors of the environment.

Myco-heterotrophy is a symbiotic relationship between certain kinds of plants and fungi, in which the plant gets all or part of its food from parasitism upon fungi rather than from photosynthesis. A myco-heterotroph is the parasitic plant partner in this relationship. Myco-heterotrophy is considered a kind of cheating relationship and myco-heterotrophs are sometimes informally referred to as "mycorrhizal cheaters". This relationship is sometimes referred to as mycotrophy, though this term is also used for plants that engage in mutualistic mycorrhizal relationships.

Rhizopogon is a genus of ectomycorrhizal basidiomycetes in the family Rhizopogonaceae. Species form hypogeous sporocarps commonly referred to as "false truffles". The general morphological characters of Rhizopogon sporocarps are a simplex or duplex peridium surrounding a loculate gleba that lacks a columnella. Basidiospores are produced upon basidia that are borne within the fungal hymenium that coats the interior surface of gleba locules. The peridium is often adorned with thick mycelial cords, also known as rhizomorphs, that attach the sporocarp to the surrounding substrate. The scientific name Rhizopogon is Greek for 'root' (Rhiz-) 'beard' (-pogon) and this name was given in reference to the rhizomorphs found on sporocarps of many species.
Hydraulic redistribution is a passive mechanism where water is transported from moist to dry soils via subterranean networks. It occurs in vascular plants that commonly have roots in both wet and dry soils, especially plants with both taproots that grow vertically down to the water table, and lateral roots that sit close to the surface. In the late 1980s, there was a movement to understand the full extent of these subterranean networks. Since then it was found that vascular plants are assisted by fungal networks which grow on the root system to promote water redistribution.
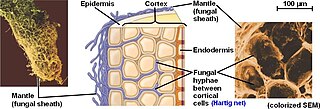
The Hartig net is the network of inward-growing hyphae, that extends into the plant host root, penetrating between plant cells in the root epidermis and cortex in ectomycorrhizal symbiosis. This network is the internal component of fungal morphology in ectomycorrhizal symbiotic structures formed with host plant roots, in addition to a hyphal mantle or sheath on the root surface, and extramatrical mycelium extending from the mantle into the surrounding soil. The Hartig net is the site of mutualistic resource exchange between the fungus and the host plant. Essential nutrients for plant growth are acquired from the soil by exploration and foraging of the extramatrical mycelium, then transported through the hyphal network across the mantle and into the Hartig net, where they are released by the fungi into the root apoplastic space for uptake by the plant. The hyphae in the Hartig net acquire sugars from the plant root, which are transported to the external mycelium to provide a carbon source to sustain fungal growth.
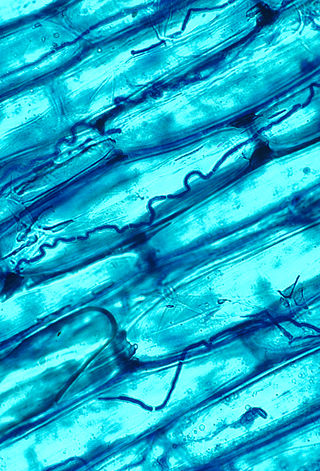
Plant use of endophytic fungi in defense occurs when endophytic fungi, which live symbiotically with the majority of plants by entering their cells, are utilized as an indirect defense against herbivores. In exchange for carbohydrate energy resources, the fungus provides benefits to the plant which can include increased water or nutrient uptake and protection from phytophagous insects, birds or mammals. Once associated, the fungi alter nutrient content of the plant and enhance or begin production of secondary metabolites. The change in chemical composition acts to deter herbivory by insects, grazing by ungulates and/or oviposition by adult insects. Endophyte-mediated defense can also be effective against pathogens and non-herbivory damage.

Soil carbon storage is an important function of terrestrial ecosystems. Soil contains more carbon than plants and the atmosphere combined. Understanding what maintains the soil carbon pool is important to understand the current distribution of carbon on Earth, and how it will respond to environmental change. While much research has been done on how plants, free-living microbial decomposers, and soil minerals affect this pool of carbon, it is recently coming to light that mycorrhizal fungi—symbiotic fungi that associate with roots of almost all living plants—may play an important role in maintaining this pool as well. Measurements of plant carbon allocation to mycorrhizal fungi have been estimated to be 5 to 20% of total plant carbon uptake, and in some ecosystems the biomass of mycorrhizal fungi can be comparable to the biomass of fine roots. Recent research has shown that mycorrhizal fungi hold 50 to 70 percent of the total carbon stored in leaf litter and soil on forested islands in Sweden. Turnover of mycorrhizal biomass into the soil carbon pool is thought to be rapid and has been shown in some ecosystems to be the dominant pathway by which living carbon enters the soil carbon pool.

An ectomycorrhiza is a form of symbiotic relationship that occurs between a fungal symbiont, or mycobiont, and the roots of various plant species. The mycobiont is often from the phyla Basidiomycota and Ascomycota, and more rarely from the Zygomycota. Ectomycorrhizas form on the roots of around 2% of plant species, usually woody plants, including species from the birch, dipterocarp, myrtle, beech, willow, pine and rose families. Research on ectomycorrhizas is increasingly important in areas such as ecosystem management and restoration, forestry and agriculture.

Ectomycorrhizal extramatrical mycelium is the collection of filamentous fungal hyphae emanating from ectomycorrhizas. It may be composed of fine, hydrophilic hypha which branches frequently to explore and exploit the soil matrix or may aggregate to form rhizomorphs; highly differentiated, hydrophobic, enduring, transport structures.

The root microbiome is the dynamic community of microorganisms associated with plant roots. Because they are rich in a variety of carbon compounds, plant roots provide unique environments for a diverse assemblage of soil microorganisms, including bacteria, fungi, and archaea. The microbial communities inside the root and in the rhizosphere are distinct from each other, and from the microbial communities of bulk soil, although there is some overlap in species composition.
Dark septate endophytes (DSE) are a group of endophytic fungi characterized by their morphology of melanized, septate, hyphae. This group is likely paraphyletic, and contain conidial as well as sterile fungi that colonize roots intracellularly or intercellularly. Very little is known about the number of fungal taxa within this group, but all are in the Ascomycota. They are found in over 600 plant species and across 114 families of angiosperms and gymnosperms and co-occur with other types of mycorrhizal fungi. They have a wide global distribution and can be more abundant in stressed environments. Much of their taxonomy, physiology, and ecology are unknown.

Suzanne Simard is a Canadian scientist and Professor in the Department of Forest and Conservation Sciences at the University of British Columbia. After growing up in the Monashee Mountains, British Columbia, she received her PhD in Forest Sciences at Oregon State University. Prior to teaching at the University of British Columbia, Simard worked as a research scientist at the British Columbia Ministry of Forests.
Plants are exposed to many stress factors such as disease, temperature changes, herbivory, injury and more. Therefore, in order to respond or be ready for any kind of physiological state, they need to develop some sort of system for their survival in the moment and/or for the future. Plant communication encompasses communication using volatile organic compounds, electrical signaling, and common mycorrhizal networks between plants and a host of other organisms such as soil microbes, other plants, animals, insects, and fungi. Plants communicate through a host of volatile organic compounds (VOCs) that can be separated into four broad categories, each the product of distinct chemical pathways: fatty acid derivatives, phenylpropanoids/benzenoids, amino acid derivatives, and terpenoids. Due to the physical/chemical constraints most VOCs are of low molecular mass, are hydrophobic, and have high vapor pressures. The responses of organisms to plant emitted VOCs varies from attracting the predator of a specific herbivore to reduce mechanical damage inflicted on the plant to the induction of chemical defenses of a neighboring plant before it is being attacked. In addition, the host of VOCs emitted varies from plant to plant, where for example, the Venus Fly Trap can emit VOCs to specifically target and attract starved prey. While these VOCs typically lead to increased resistance to herbivory in neighboring plants, there is no clear benefit to the emitting plant in helping nearby plants. As such, whether neighboring plants have evolved the capability to "eavesdrop" or whether there is an unknown tradeoff occurring is subject to much scientific debate. As related to the aspect of meaning-making, the field is also identified as phytosemiotics.
Orchid mycorrhizae are endomycorrhizal fungi which develop symbiotic relationships with the roots and seeds of plants of the family Orchidaceae. Nearly all orchids are myco-heterotrophic at some point in their life cycle. Orchid mycorrhizae are critically important during orchid germination, as an orchid seed has virtually no energy reserve and obtains its carbon from the fungal symbiont.
Mycorrhizal amelioration of heavy metals or pollutants is a process by which mycorrhizal fungi in a mutualistic relationship with plants can sequester toxic compounds from the environment, as a form of bioremediation.

Rhizopogon salebrosus is a mushroom species within the Rhizopogon sub-genus Amylopogon. R.salebrosus is a monotropoid mycorrhiza that is of vital importance to the ecology of conifer forests, especially in the Pacific Northwest region of North America. Although it is native to North America, R. salebrosus has been found in Europe and its range is generally limited to mountainous regions with sufficient precipitation. The mycoheterotrophic plant, Pterospora andromedea is often found in an obligate association with R. salebrosus in western parts of the U.S. Eastern populations of P. andromedea are typically symbiotic with another Rhizopogon sub species, R. kretzerae.


