Related Research Articles
In biology, kingdom is the second highest taxonomic rank, just below domain. Kingdoms are divided into smaller groups called phyla. Traditionally, some textbooks from the United States and Canada used a system of six kingdoms while textbooks in Great Britain, India, Greece, Brazil and other countries use five kingdoms only. Some recent classifications based on modern cladistics have explicitly abandoned the term kingdom, noting that the traditional kingdoms are not monophyletic, meaning that they do not consist of all the descendants of a common ancestor.

Euryarchaeota is a phylum of archaea. It is one of two phyla of archaea, the other being crenarchaeota. Euryarchaeota are highly diverse and include methanogens, which produce methane and are often found in intestines, halobacteria, which survive extreme concentrations of salt, and some extremely thermophilic aerobes and anaerobes, which generally live at temperatures between 41 and 122 °C. They are separated from the other archaeans based mainly on rRNA sequences and their unique DNA polymerase.
The Aquificae phylum is a diverse collection of bacteria that live in harsh environmental settings. The name 'Aquificae' was given to this phylum based on an early genus identified within this group, Aquifex, which is able to produce water by oxidizing hydrogen. They have been found in springs, pools, and oceans. They are autotrophs, and are the primary carbon fixers in their environments. These bacteria are Gram-negative, non-spore-forming rods. They are true bacteria as opposed to the other inhabitants of extreme environments, the Archaea.

Deinococcus–Thermus is a phylum of bacteria that are highly resistant to environmental hazards, also known as extremophiles. These bacteria have thick cell walls that give them gram-positive stains, but they include a second membrane and so are closer in structure to those of gram-negative bacteria. Cavalier-Smith calls this clade Hadobacteria.
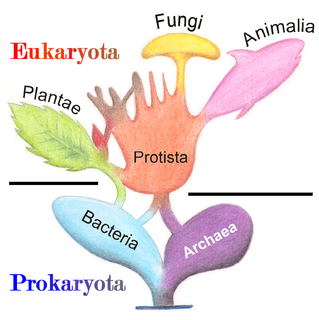
The two-empire system was the top-level biological classification system in general use before the establishment of the three-domain system. It classified cellular life into Prokaryota and Eukaryota as either "empires" or "superkingdoms". When the three-domain system was introduced, some biologists preferred the two-superkingdom system, claiming that the three-domain system overemphasized the division between Archaea and Bacteria. However, given the current state of knowledge and the rapid progress in biological scientific advancement, especially due to genetic analyses, that view has all but vanished.
Neomura is a possible clade composed of the two domains of life of Archaea and Eukaryota. The group was named by Thomas Cavalier-Smith in 2002. Its name means "new walls", reflecting his hypothesis that it evolved from Bacteria, and one of the major changes was the replacement of peptidoglycan cell walls with other glycoproteins. As of August 2017, the neomuran hypothesis is not accepted by most workers; molecular phylogenies suggest that eukaryotes are most closely related to one group of archaeans and evolved from them, rather than forming a clade with all archaeans.

Gracilicutes is a clade in bacterial phylogeny.

The PVC group is a superphylum of bacteria named after its three important members, Planctomycetes, Verrucomicrobia, and Chlamydiae. Cavalier-Smith postulated that the PVC bacteria probably lost or reduced their peptidoglycan cell wall twice. It has been hypothesised that a member of the PVC clade might have been the host cell in the endosymbiotic event that gave rise to the first proto-eukaryotic cell.

In the taxonomy of microorganisms, the Methanomicrobia are a class of the Euryarchaeota.
Archaea constitute a domain of single-celled organisms. These microorganisms lack cell nuclei and are therefore prokaryotes. Archaea were initially classified as bacteria, receiving the name archaebacteria, but this term has fallen out of use.

Terrabacteria is a taxon containing approximately two-thirds of prokaryote species, including those in the gram positive phyla as well as the phyla Cyanobacteria, Chloroflexi, and Deinococcus-Thermus.
Bacterial taxonomy is the taxonomy, i.e. the rank-based classification, of bacteria.
There are several models of the Branching order of bacterial phyla, one of these was proposed in 1987 paper by Carl Woese.
There are several models of the Branching order of bacterial phyla, one of these was proposed in 2006 by Ciccarelli et al. for their iTOL project.
There are several models of the Branching order of bacterial phyla, one of these was proposed in 2004 by Battistuzzi and Hedges, note the coinage of the taxa Terrabacteria and Hydrobacteria.
There are several models of the Branching order of bacterial phyla, one of these was proposed in 2001 by Gupta based on conserved indels or protein, termed "protein signatures", an alternative approach to molecular phylogeny. Some problematic exceptions and conflicts are present to these conserved indels, however, they are in agreement with several groupings of classes and phyla. One feature of the cladogram obtained with this method is the clustering of cell wall morphology from monoderms to transitional diderms to traditional diderms.
Conserved signature inserts and deletions (CSIs) in protein sequences provide an important category of molecular markers for understanding phylogenetic relationships. CSIs, brought about by rare genetic changes, provide useful phylogenetic markers that are generally of defined size and they are flanked on both sides by conserved regions to ensure their reliability. While indels can be arbitrary inserts or deletions, CSIs are defined as only those protein indels that are present within conserved regions of the protein.
The Scotokaryotes (Cavalier-Smith) is a proposed basal Neokaryote clade as sister of the Diaphoretickes. Basal Scotokaryote groupings are the Metamonads, the Malawimonas and the Podiata. In this phylogeny the Discoba are sometimes seen as paraphyletic and basal Eukaryotes.
Bacterial evolution may refer to the biological evolution of bacteria as studied in:
There are several models of the branching order of bacterial phyla, one of these is the Genome Taxonomy Database (GTDB).
References
- 1 2 Cavalier-Smith, T (2002). "The neomuran origin of archaebacteria, the negibacterial root of the universal tree and bacterial megaclassification". International Journal of Systematic and Evolutionary Microbiology. 52 (Pt 1): 7–76. doi: 10.1099/00207713-52-1-7 . PMID 11837318.
- 1 2 Cavalier-Smith T (2006). "Rooting the tree of life by transition analyses". Biol. Direct. 1: 19. doi:10.1186/1745-6150-1-19. PMC 1586193 . PMID 16834776. Archived from the original on 18 December 2019. Retrieved 21 November 2011.
| Medical microbiology | |||||||
|---|---|---|---|---|---|---|---|
| Biochemistry and ecology |
| ||||||
| Shape | |||||||
| Structure |
| ||||||
| Taxonomy and evolution | |||||||