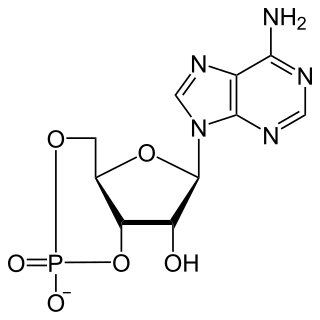
Cyclic adenosine monophosphate is a second messenger, or cellular signal occurring within cells, that is important in many biological processes. cAMP is a derivative of adenosine triphosphate (ATP) and used for intracellular signal transduction in many different organisms, conveying the cAMP-dependent pathway.

Cholera is an infection of the small intestine by some strains of the bacterium Vibrio cholerae. Symptoms may range from none, to mild, to severe. The classic symptom is large amounts of watery diarrhea lasting a few days. Vomiting and muscle cramps may also occur. Diarrhea can be so severe that it leads within hours to severe dehydration and electrolyte imbalance. This may result in sunken eyes, cold skin, decreased skin elasticity, and wrinkling of the hands and feet. Dehydration can cause the skin to turn bluish. Symptoms start two hours to five days after exposure.

Vibrio cholerae is a species of Gram-negative, facultative anaerobe and comma-shaped bacteria. The bacteria naturally live in brackish or saltwater where they attach themselves easily to the chitin-containing shells of crabs, shrimp, and other shellfish. Some strains of V. cholerae are pathogenic to humans and cause a deadly disease called cholera, which can be derived from the consumption of undercooked or raw marine life species or drinking contaminated water.
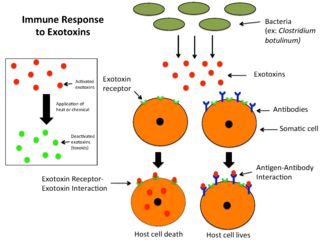
An exotoxin is a toxin secreted by bacteria. An exotoxin can cause damage to the host by destroying cells or disrupting normal cellular metabolism. They are highly potent and can cause major damage to the host. Exotoxins may be secreted, or, similar to endotoxins, may be released during lysis of the cell. Gram negative pathogens may secrete outer membrane vesicles containing lipopolysaccharide endotoxin and some virulence proteins in the bounding membrane along with some other toxins as intra-vesicular contents, thus adding a previously unforeseen dimension to the well-known eukaryote process of membrane vesicle trafficking, which is quite active at the host–pathogen interface.

An enterotoxin is a protein exotoxin released by a microorganism that targets the intestines. They can be chromosomally or plasmid encoded. They are heat labile (>60⁰), of low molecular weight and water-soluble. Enterotoxins are frequently cytotoxic and kill cells by altering the apical membrane permeability of the mucosal (epithelial) cells of the intestinal wall. They are mostly pore-forming toxins, secreted by bacteria, that assemble to form pores in cell membranes. This causes the cells to die.

ADP-ribosylation factor 6 (ARF6) is a member of the ADP ribosylation factor family of GTP-binding proteins. ARF6 has a variety of cellular functions that are frequently involved in trafficking of biological membranes and transmembrane protein cargo. ARF6 has specifically been implicated in endocytosis of plasma membrane proteins and also, to a lesser extent, plasma membrane protein recycling.

ADP ribosylation factors (ARFs) are members of the ARF family of GTP-binding proteins of the Ras superfamily. ARF family proteins are ubiquitous in eukaryotic cells, and six highly conserved members of the family have been identified in mammalian cells. Although ARFs are soluble, they generally associate with membranes because of N-terminus myristoylation. They function as regulators of vesicular traffic and actin remodelling.

GM1 (monosialotetrahexosylganglioside) the "prototype" ganglioside, is a member of the ganglio series of gangliosides which contain one sialic acid residue. GM1 has important physiological properties and impacts neuronal plasticity and repair mechanisms, and the release of neurotrophins in the brain. Besides its function in the physiology of the brain, GM1 acts as the site of binding for both cholera toxin and E. coli heat-labile enterotoxin.

Pore-forming proteins are usually produced by bacteria, and include a number of protein exotoxins but may also be produced by other organisms such as apple snails that produce perivitellin-2 or earthworms, who produce lysenin. They are frequently cytotoxic, as they create unregulated pores in the membrane of targeted cells.
The AB5 toxins are six-component protein complexes secreted by certain pathogenic bacteria known to cause human diseases such as cholera, dysentery, and hemolytic–uremic syndrome. One component is known as the A subunit, and the remaining five components are B subunits. All of these toxins share a similar structure and mechanism for entering targeted host cells. The B subunit is responsible for binding to receptors to open up a pathway for the A subunit to enter the cell. The A subunit is then able to use its catalytic machinery to take over the host cell's regular functions.

ADP-ribosylation is the addition of one or more ADP-ribose moieties to a protein. It is a reversible post-translational modification that is involved in many cellular processes, including cell signaling, DNA repair, gene regulation and apoptosis. Improper ADP-ribosylation has been implicated in some forms of cancer. It is also the basis for the toxicity of bacterial compounds such as cholera toxin, diphtheria toxin, and others.

ADP-ribosylation factor 1 is a protein that in humans is encoded by the ARF1 gene.

Sambhunath De ; was an Indian medical scientist and researcher, who discovered the cholera toxin, the animal model of cholera, and successfully demonstrated the method of transmission of cholera pathogen Vibrio cholerae.
Microbial toxins are toxins produced by micro-organisms, including bacteria, fungi, protozoa, dinoflagellates, and viruses. Many microbial toxins promote infection and disease by directly damaging host tissues and by disabling the immune system. Endotoxins most commonly refer to the lipopolysaccharide (LPS) or lipooligosaccharide (LOS) that are in the outer plasma membrane of Gram-negative bacteria. The botulinum toxin, which is primarily produced by Clostridium botulinum and less frequently by other Clostridium species, is the most toxic substance known in the world. However, microbial toxins also have important uses in medical science and research. Currently, new methods of detecting bacterial toxins are being developed to better isolate and understand these toxins. Potential applications of toxin research include combating microbial virulence, the development of novel anticancer drugs and other medicines, and the use of toxins as tools in neurobiology and cellular biology.
In molecular biology, the heat-labile enterotoxin family includes Escherichia coli heat-labile enterotoxin and cholera toxin (Ctx) secreted by Vibrio cholerae.
The CTXφ bacteriophage is a filamentous bacteriophage. It is a positive-strand DNA virus with single-stranded DNA (ssDNA).
Vibrio campbellii is a Gram-negative, curved rod-shaped, marine bacterium closely related to its sister species, Vibrio harveyi. It is an emerging pathogen in aquatic organisms.
John Mekalanos is a microbiologist who is primarily known for leading one of the first teams that reported the discovery of the type VI secretion system as well as his work on the pathogenicity of the bacterial species Vibrio cholerae, its toxin, and its secretion systems. Since 1998, he has been a member of the National Academy of Sciences.
Jan Roland Holmgren is a Swedish physician, microbiologist, immunologist, and vaccinologist, known for his research on cholera and mucosal immunology, specifically, for his leadership in developing "the world's first effective oral cholera vaccine".
Clathrin-independent endocytosis refers to the cellular process by which cells internalize extracellular molecules and particles through mechanisms that do not rely on the protein clathrin, playing a crucial role in diverse physiological processes such as nutrient uptake, membrane turnover, and cellular signaling.


