An anapsid is an amniote whose skull lacks one or more skull openings near the temples. Traditionally, the Anapsida are the most primitive subclass of amniotes, the ancestral stock from which Synapsida and Diapsida evolved, making anapsids paraphyletic. It is however doubtful that all anapsids lack temporal fenestra as a primitive trait, and that all the groups traditionally seen as anapsids truly lacked fenestra.

Diapsids are a clade of sauropsids, distinguished from more primitive eureptiles by the presence of two holes, known as temporal fenestrae, in each side of their skulls. The group first appeared about three hundred million years ago during the late Carboniferous period. All diapsids other than the most primitive ones in the clade Araeoscelidia are sometimes placed into the clade Neodiapsida. The diapsids are extremely diverse, and include birds and all modern reptile groups, including turtles, which were historically thought to lie outside the group. Although some diapsids have lost either one hole (lizards), or both holes, or have a heavily restructured skull, they are still classified as diapsids based on their ancestry. At least 17,084 species of diapsid animals are extant: 9,159 birds, and 7,925 snakes, lizards, tuatara, turtles, and crocodiles.

Sauropsida is a clade of amniotes, broadly equivalent to the class Reptilia, though typically used in a broader sense to include both extinct stem-group relatives of modern reptiles, as well as birds. The most popular definition states that Sauropsida is the sibling taxon to Synapsida, the other clade of amniotes which includes mammals as its only modern representatives. Although early synapsids have historically been referred to as "mammal-like reptiles", all synapsids are more closely related to mammals than to any modern reptile. Sauropsids, on the other hand, include all amniotes more closely related to modern reptiles than to mammals. This includes Aves (birds), which are now recognized as a subgroup of archosaurian reptiles despite originally being named as a separate class in Linnaean taxonomy.
Several groups of tetrapods have undergone secondary aquatic adaptation, an evolutionary transition from being purely terrestrial to living at least part of the time in water. These animals are called "secondarily aquatic" because although their ancestors lived on land for hundreds of millions of years, they all originally descended from aquatic animals. These ancestral tetrapods had never left the water, and were thus primarily aquatic, like modern fishes. Secondary aquatic adaptations tend to develop in early speciation as the animal ventures into water in order to find available food. As successive generations spend more time in the water, natural selection causes the acquisition of more adaptations. Animals of later generations may spend most their life in the water, coming ashore for mating. Finally, fully adapted animals may take to mating and birthing in water or ice.

Marine reptiles are reptiles which have become secondarily adapted for an aquatic or semiaquatic life in a marine environment.

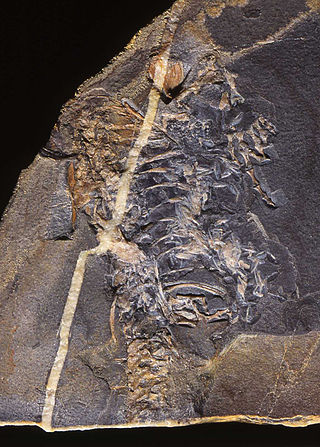
Mesosaurs were a group of small aquatic reptiles that lived during the early Permian period (Cisuralian), roughly 299 to 270 million years ago. Mesosaurs were the first known aquatic reptiles, having apparently returned to an aquatic lifestyle from more terrestrial ancestors. It is uncertain which and how many terrestrial traits these ancestors displayed; recent research cannot establish with confidence if the first amniotes were fully terrestrial, or only amphibious. Most authors consider mesosaurs to have been aquatic, although adult animals may have been amphibious, rather than completely aquatic, as indicated by their moderate skeletal adaptations to a semiaquatic lifestyle. Similarly, their affinities are uncertain; they may have been among the most basal sauropsids or among the most basal parareptiles.

"Labyrinthodontia" is an informal grouping of extinct predatory amphibians which were major components of ecosystems in the late Paleozoic and early Mesozoic eras. Traditionally considered a subclass of the class Amphibia, modern classification systems recognize that labyrinthodonts are not a formal natural group (clade) exclusive of other tetrapods. Instead, they consistute an evolutionary grade, ancestral to living tetrapods such as lissamphibians and amniotes. "Labyrinthodont"-grade vertebrates evolved from lobe-finned fishes in the Devonian, though a formal boundary between fish and amphibian is difficult to define at this point in time.

Friedrich von Huene, born Friedrich Richard von Hoinigen, was a German paleontologist who renamed more dinosaurs in the early 20th century than anyone else in Europe. He also made key contributions about various Permo-Carboniferous limbed vertebrates.

Reptiliomorpha is a clade containing the amniotes and those tetrapods that share a more recent common ancestor with amniotes than with living amphibians (lissamphibians). It was defined by Michel Laurin (2001) and Vallin and Laurin (2004) as the largest clade that includes Homo sapiens, but not Ascaphus truei. Laurin and Reisz (2020) defined Pan-Amniota as the largest total clade containing Homo sapiens, but not Pipa pipa, Caecilia tentaculata, and Siren lacertina.

Diadectomorpha is a clade of large tetrapods that lived in Euramerica during the Carboniferous and Early Permian periods and in Asia during Late Permian (Wuchiapingian), They have typically been classified as advanced reptiliomorphs positioned close to, but outside of the clade Amniota, though some recent research has recovered them as the sister group to the traditional Synapsida within Amniota, based on inner ear anatomy and cladistic analyses. They include both large carnivorous and even larger herbivorous forms, some semi-aquatic and others fully terrestrial. The diadectomorphs seem to have originated during late Mississippian times, although they only became common after the Carboniferous rainforest collapse and flourished during the Late Pennsylvanian and Early Permian periods.
Varanopidae is an extinct family of amniotes that resembled monitor lizards and may have filled a similar niche, hence the name. Typically, they are considered synapsids that evolved from an Archaeothyris-like synapsid in the Late Carboniferous. However, some recent studies have recovered them being taxonomically closer to diapsid reptiles. A varanopid from the latest Middle Permian Pristerognathus Assemblage Zone is the youngest known varanopid and the last member of the "pelycosaur" group of synapsids.

Ophiacodon is an extinct genus of synapsid belonging to the family Ophiacodontidae that lived from the Late Carboniferous to the Early Permian in North America and Europe. The genus was named along with its type species O. mirus by paleontologist Othniel Charles Marsh in 1878 and currently includes five other species. As an ophiacodontid, Ophiacodon is one of the most basal synapsids and is close to the evolutionary line leading to mammals.

Parareptilia ("near-reptiles") is a subclass or clade of basal sauropsids/reptiles, typically considered the sister taxon to Eureptilia. Parareptiles first arose near the end of the Carboniferous period and achieved their highest diversity during the Permian period. Several ecological innovations were first accomplished by parareptiles among reptiles. These include the first reptiles to return to marine ecosystems (mesosaurs), the first bipedal reptiles, the first reptiles with advanced hearing systems, and the first large herbivorous reptiles. The only parareptiles to survive into the Triassic period were the procolophonoids, a group of small generalists, omnivores, and herbivores. The largest family of procolophonoids, the procolophonids, rediversified in the Triassic, but subsequently declined and became extinct by the end of the period.

Claudiosaurus is an extinct genus of diapsid reptiles from the Late Permian Sakamena Formation of the Morondava Basin, Madagascar. It has been suggested to be semi-aquatic.

Stereosternum tumidum is an extinct genus of mesosaur marine reptile from the Early Permian of Brazil and also the Great Karoo Basin of South Africa. The taxon mesosaur is a monophyletic group containing Brazilosaurus sanpauloensis and Mesosaurus tenuidens.

Captorhinus is an extinct genus of captorhinid reptiles that lived during the Permian period. Its remains are known from North America and possibly South America.
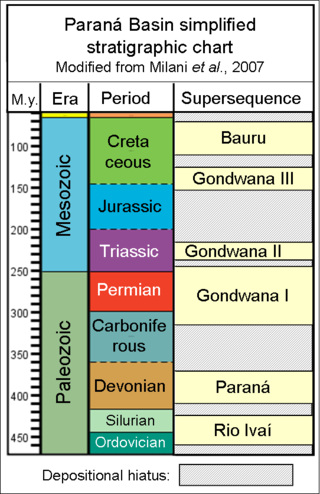
Irati Formation is the name of a geological formation of the Paraná Basin in Brazil. It has previously been dated as Late Permian using palynomorphs, but is now dated as Early Permian using zircon ages obtained from bentonite layers. The base of the formation has been dated at 278.4 ± 2.2 Ma. Exposures of the Irati Formation are to be found in the South, southeastern Brazil and in the states of Goiás, Mato Grosso, São Paulo, Paraná, Santa Catarina, Rio Grande do Sul and Mato Grosso do Sul. The formation is part of the Passa Dios Group, underlying the Serra Alta Formation and overlying the Palermo Formation. The formation has been deposited in a restricted marine environment. The Irati Formation, with a maximum thickness of 80 metres (260 ft), was defined and named by White in 1908.
Reptiles arose about 320 million years ago during the Carboniferous period. Reptiles, in the traditional sense of the term, are defined as animals that have scales or scutes, lay land-based hard-shelled eggs, and possess ectothermic metabolisms. So defined, the group is paraphyletic, excluding endothermic animals like birds that are descended from early traditionally-defined reptiles. A definition in accordance with phylogenetic nomenclature, which rejects paraphyletic groups, includes birds while excluding mammals and their synapsid ancestors. So defined, Reptilia is identical to Sauropsida.

The Mangrullo Formation is an Early Permian (Artinskian) fossiliferous geological formation in northeastern Uruguay. Some authors alternatively group it together with the Paso Aguiar Formation and the Frayle Muerto Formation as the three subdivisions of the Melo Formation, in which case it is referred to as the Mangrullo Member. Like the correlated formations of Irati and Whitehill, it is known for its abundant mesosaur fossils. It also contains the oldest known Konservat-Lagerstätte in South America, as well as the oldest known fossils of amniote embryos.
Graciela Helena Piñeiro Martínez is a Uruguayan biologist and paleontologist.