
Mesosaurs were a group of small aquatic reptiles that lived during the early Permian period (Cisuralian), roughly 299 to 270 million years ago. Mesosaurs were the first known aquatic reptiles, having apparently returned to an aquatic lifestyle from more terrestrial ancestors. It is uncertain which and how many terrestrial traits these ancestors displayed; recent research cannot establish with confidence if the first amniotes were fully terrestrial, or only amphibious. Most authors consider mesosaurs to have been aquatic, although adult animals may have been amphibious, rather than completely aquatic, as indicated by their moderate skeletal adaptations to a semiaquatic lifestyle. Similarly, their affinities are uncertain; they may have been among the most basal sauropsids or among the most basal parareptiles.

Neodiapsida is a clade, or major branch, of the reptilian family tree, typically defined as including all diapsids apart from some early primitive types known as the araeoscelidians. Modern reptiles and birds belong to the neodiapsid subclade Sauria.

Youngina is an extinct genus of diapsid reptile from the Late Permian Beaufort Group of the Karoo Red Beds of South Africa. This, and a few related forms, make up the family Younginidae, within the order Eosuchia. Eosuchia, having become a wastebasket taxon for many probably distantly-related primitive diapsid reptiles ranging from the Late Carboniferous to the Eocene, Romer proposed that it be replaced by Younginiformes.

Petrolacosaurus is an extinct genus of diapsid reptile from the late Carboniferous period. It was a small, 40-centimetre (16 in) long reptile, and one of the earliest known reptile with two temporal fenestrae. This means that it was at the base of Diapsida, the largest and most successful radiation of reptiles that would eventually include all modern reptile groups, as well as dinosaurs and other famous extinct reptiles such as plesiosaurs, ichthyosaurs, and pterosaurs. However, Petrolacosaurus itself was part of Araeoscelida, a short-lived early branch of the diapsid family tree which went extinct in the mid-Permian.

Coelurosauravus is an extinct genus of gliding reptile, known from the Late Permian of Madagascar. Like other members of the family Weigeltisauridae, members of this genus possessed long, rod-like ossifications projecting outwards from the body. These bony rods were not extensions of the ribs but were instead a feature unique to weigeltisaurids. It is believed that during life, these structures formed folding wings used for gliding flight, similar to living gliding Draco lizards.

Parareptilia ("near-reptiles") is an extinct subclass or clade of basal sauropsids/reptiles, typically considered the sister taxon to Eureptilia. Parareptiles first arose near the end of the Carboniferous period and achieved their highest diversity during the Permian period. Several ecological innovations were first accomplished by parareptiles among reptiles. These include the first reptiles to return to marine ecosystems (mesosaurs), the first bipedal reptiles, the first reptiles with advanced hearing systems, and the first large herbivorous reptiles. The only parareptiles to survive into the Triassic period were the procolophonoids, a group of small generalists, omnivores, and herbivores. The largest family of procolophonoids, the procolophonids, rediversified in the Triassic, but subsequently declined and became extinct by the end of the period.

Eosuchians are an extinct order of diapsid reptiles. Depending on which taxa are included the order may have ranged from the late Carboniferous to the Eocene but the consensus is that eosuchians are confined to the Permian and Triassic.

Younginiformes is a group of diapsid reptiles known from the Permian-Triassic of Africa and Madagascar. It has been used as a replacement for "Eosuchia". Younginiformes were historically suggested to be lepidosauromorphs, but were later suggested to be basal non-saurian neodiapsids. The group is sometimes divided into two families, Tangasauridae and Younginidae. The monophyly of the group is disputed. A 2009 study found them to be an unresolved polytomy at the base of Neodiapsida, while a 2011 study recovered the group as paraphyletic. A 2022 study recovered the Younginiformes as a monophyletic group of basal neodiapsid reptiles, also including Claudiosaurus and Saurosternon as part of the group. Some younginiforms like Hovasaurus and Acerosodontosaurus are thought to have had an amphibious lifestyle, while others like Kenyasaurus, Thadeosaurus and Youngina were probably terrestrial.

Claudiosaurus is an extinct genus of diapsid reptiles from the Late Permian Sakamena Formation of the Morondava Basin, Madagascar. It has been suggested to be semi-aquatic.

Thadeosaurus is an extinct genus of diapsid reptile belonging to the family Younginidae. Fossils have been found in the Lower Sakamena Formation of the Morondava Basin, Madagascar in 1981, and date to the late Permian to the early Triassic period.
Hovasaurus is an extinct genus of basal diapsid reptile. It lived in what is now Madagascar during the Late Permian and Early Triassic, being a survivor of the Permian–Triassic extinction event and the paleontologically youngest member of the Tangasauridae. Fossils have been found in the Permian Lower and Triassic Middle Sakamena Formations of the Sakamena Group, where it is amongst the commonest fossils. Its morphology suggests an aquatic ecology.

Araeoscelidia or Araeoscelida is a clade of extinct diapsid reptiles superficially resembling lizards, extending from the Late Carboniferous to the Early Permian. The group contains the genera Araeoscelis, Petrolacosaurus, the possibly aquatic Spinoaequalis, and less well-known genera such as Kadaliosaurus and Zarcasaurus. This clade is usually considered to be the sister group to all later diapsids.

Acerosodontosaurus is an extinct genus of neodiapsid reptiles that lived during the Late Permian of Madagascar. The only species of Acerosodontosaurus, A. piveteaui, is known from a natural mold of a single partial skeleton including a crushed skull and part of the body and limbs. The fossil was discovered in deposits of the Lower Sakamena Formation. Based on skeletal characteristics, it has been suggested that Acerosodontosaurus individuals were at least partially aquatic.
Tangasauridae is an extinct family of diapsids known from fossil specimens from Madagascar, Kenya and Tanzania that are Late Permian to Early Triassic in age. Fossils have been found of numerous specimens of common members of this family such as Hovasaurus in different stages of ontogenic development. Recent material from the Middle Sakamena Formation of the Morondava Basin of Madagascar that dates back to the early Triassic period suggests that the Tangasauridae were relatively unaffected by the Permian-Triassic extinction event.

Ianthodon is an extinct genus of basal haptodontiform synapsids from the Late Carboniferous about 304 million years ago. The taxon was discovered and named by Kissel & Reisz in 2004. The only species in the taxon, Ianthodon schultzei, was found by separating it from a block that also contained the remains of Petrolacosaurus and Haptodus. The evolutionary significance of the taxon wasn't realized until a publication in 2015. The fossil of this organism was discovered in Garnett, Kansas.

Weigeltisauridae is a family of gliding neodiapsid reptiles that lived during the Late Permian, between 259.51 and 251.9 million years ago. Fossils of weigeltisaurids have been found in Madagascar, Germany, Great Britain, and Russia. They are characterized by long, hollow rod-shaped bones extending from the torso that probably supported wing-like membranes. Similar membranes are also found in several other extinct reptiles such as kuehneosaurids and Mecistotrachelos, as well as living gliding lizards, although each group evolved these structures independently.
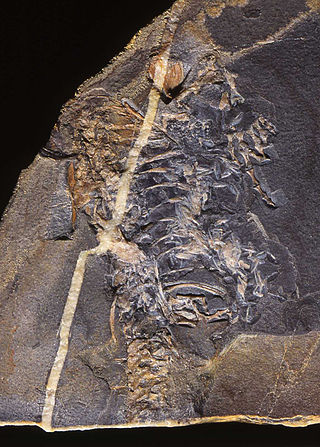
Reptiles arose about 320 million years ago during the Carboniferous period. Reptiles, in the traditional sense of the term, are defined as animals that have scales or scutes, lay land-based hard-shelled eggs, and possess ectothermic metabolisms. So defined, the group is paraphyletic, excluding endothermic animals like birds that are descended from early traditionally-defined reptiles. A definition in accordance with phylogenetic nomenclature, which rejects paraphyletic groups, includes birds while excluding mammals and their synapsid ancestors. So defined, Reptilia is identical to Sauropsida.
Kenyasaurus is an extinct genus of basal tangasaurid known from the Early Triassic period of Coast Province, southeastern Kenya. It contains a single species, Kenyasaurus mariakaniensis.
Younginidae is an extinct family of diapsid reptiles from the Late Permian and Early Triassic. In a phylogenetic context, younginids are near the base of the clade Neodiapsida. Younginidae includes the species Youngina capensis from the Late Permian of South Africa and Thadeosaurus colcanapi from the Late Permian and Early Triassic of Madagascar. Heleosuchus griesbachi from the Late Permian of South Africa may also be a member of the family.

Aenigmastropheus is an extinct genus of early archosauromorph reptiles known from the middle Late Permian Usili Formation of Songea District, southern Tanzania. It contains a single species, Aenigmastropheus parringtoni, known solely from UMZC T836, a partial postcranial skeleton of a mature individual. It was collected in 1933, and first described in 1956, as a "problematic reptile" due to its unique morphology. Therefore, a binomial name was erected for this specimen in 2014. Aenigmastropheus was probably fully terrestrial.






