An anapsid is an amniote whose skull lacks one or more skull openings near the temples. Traditionally, the Anapsida are the most primitive subclass of amniotes, the ancestral stock from which Synapsida and Diapsida evolved, making anapsids paraphyletic. It is however doubtful that all anapsids lack temporal fenestra as a primitive trait, and that all the groups traditionally seen as anapsids truly lacked fenestra.

Sauria is the clade containing the most recent common ancestor of Archosauria and Lepidosauria, and all its descendants. Since most molecular phylogenies recover turtles as more closely related to archosaurs than to lepidosaurs as part of Archelosauria, Sauria can be considered the crown group of diapsids, or reptiles in general. Depending on the systematics, Sauria includes all modern reptiles or most of them as well as various extinct groups.

Diapsids are a clade of sauropsids, distinguished from more primitive eureptiles by the presence of two holes, known as temporal fenestrae, in each side of their skulls. The group first appeared about three hundred million years ago during the late Carboniferous period. All diapsids other than the most primitive ones in the clade Araeoscelidia are sometimes placed into the clade Neodiapsida. The diapsids are extremely diverse, and include birds and all modern reptile groups, including turtles, which were historically thought to lie outside the group. Although some diapsids have lost either one hole (lizards), or both holes, or have a heavily restructured skull, they are still classified as diapsids based on their ancestry. At least 17,084 species of diapsid animals are extant: 9,159 birds, and 7,925 snakes, lizards, tuatara, turtles, and crocodiles.

Mesosaurs were a group of small aquatic reptiles that lived during the early Permian period (Cisuralian), roughly 299 to 270 million years ago. Mesosaurs were the first known aquatic reptiles, having apparently returned to an aquatic lifestyle from more terrestrial ancestors. It is uncertain which and how many terrestrial traits these ancestors displayed; recent research cannot establish with confidence if the first amniotes were fully terrestrial, or only amphibious. Most authors consider mesosaurs to have been aquatic, although adult animals may have been amphibious, rather than completely aquatic, as indicated by their moderate skeletal adaptations to a semiaquatic lifestyle. Similarly, their affinities are uncertain; they may have been among the most basal sauropsids or among the most basal parareptiles.

Archosauromorpha is a clade of diapsid reptiles containing all reptiles more closely related to archosaurs rather than lepidosaurs. Archosauromorphs first appeared during the late Middle Permian or Late Permian, though they became much more common and diverse during the Triassic period.

Avicephala is a potentially polyphyletic grouping of extinct diapsid reptiles that lived during the Late Permian and Triassic periods characterised by superficially bird-like skulls and arboreal lifestyles. As a clade, Avicephala is defined as including the gliding weigeltisaurids and the arboreal drepanosaurs to the exclusion of other major diapsid groups. This relationship is not recovered in the majority of phylogenetic analyses of early diapsids and so Avicephala is typically regarded as an unnatural grouping. However, the clade was recovered again in 2021 in a redescription of Weigeltisaurus, raising the possibility that the clade may be valid after all.

Parareptilia ("near-reptiles") is a subclass or clade of basal sauropsids/reptiles, typically considered the sister taxon to Eureptilia. Parareptiles first arose near the end of the Carboniferous period and achieved their highest diversity during the Permian period. Several ecological innovations were first accomplished by parareptiles among reptiles. These include the first reptiles to return to marine ecosystems (mesosaurs), the first bipedal reptiles, the first reptiles with advanced hearing systems, and the first large herbivorous reptiles. The only parareptiles to survive into the Triassic period were the procolophonoids, a group of small generalists, omnivores, and herbivores. The largest family of procolophonoids, the procolophonids, rediversified in the Triassic, but subsequently declined and became extinct by the end of the period.

Younginiformes is a group of diapsid reptiles known from the Permian-Triassic of Africa and Madagascar. It has been used as a replacement for "Eosuchia". Younginiformes were historically suggested to be lepidosauromorphs, but were later suggested to be basal non-saurian neodiapsids. The group is sometimes divided into two families, Tangasauridae and Younginidae. The monophyly of the group is disputed. A 2009 study found them to be an unresolved polytomy at the base of Neodiapsida, while a 2011 study recovered the group as paraphyletic. A 2022 study recovered the Younginiformes as a monophyletic group of basal neodiapsid reptiles, also including Claudiosaurus and Saurosternon as part of the group. Some younginiforms like Hovasaurus and Acerosodontosaurus are thought to have had an amphibious lifestyle, while others like Kenyasaurus, Thadeosaurus and Youngina were probably terrestrial.

Claudiosaurus is an extinct genus of diapsid reptiles from the Late Permian Sakamena Formation of the Morondava Basin, Madagascar. It has been suggested to be semi-aquatic.

Hovasaurus is an extinct genus of basal diapsid reptile. It lived in what is now Madagascar during the Late Permian and Early Triassic, being a survivor of the Permian–Triassic extinction event and the paleontologically youngest member of the Tangasauridae. Fossils have been found in the Permian Lower and Triassic Middle Sakamena Formations of the Sakamena Group, where it is amongst the commonest fossils. Its morphology suggests an aquatic ecology.
Araeoscelidia or Araeoscelida is a clade of extinct diapsid reptiles superficially resembling lizards, extending from the Late Carboniferous to the Early Permian. The group contains the genera Araeoscelis, Petrolacosaurus, the possibly aquatic Spinoaequalis, and less well-known genera such as Kadaliosaurus and Zarcasaurus. This clade is usually considered to be the sister group to all later diapsids.

Drepanosaurs are a group of extinct reptiles that lived between the Carnian and Rhaetian stages of the late Triassic Period, approximately between 230 and 210 million years ago. The various species of drepanosaurid were characterized by specialized grasping limbs and often prehensile tails, adaptions for arboreal (tree-dwelling) and fossorial (digging) lifestyles, with some having also been suggested to be aquatic. Fossils of drepanosaurs have been found in Arizona, New Mexico, New Jersey, Utah, England, and northern Italy. The name is taken from the family's namesake genus Drepanosaurus, which means "sickle lizard," a reference to their strongly curved claws.

Acerosodontosaurus is an extinct genus of neodiapsid reptiles that lived during the Late Permian of Madagascar. The only species of Acerosodontosaurus, A. piveteaui, is known from a natural mold of a single partial skeleton including a crushed skull and part of the body and limbs. The fossil was discovered in deposits of the Lower Sakamena Formation. Based on skeletal characteristics, it has been suggested that Acerosodontosaurus individuals were at least partially aquatic.

Eunotosaurus is an extinct genus of amniote, possibly a close relative of turtles. Eunotosaurus lived in the late Middle Permian and fossils can be found in the Karoo Supergroup of South Africa. Eunotosaurus resided in the swamps of southern Africa. Its ribs were wide and flat, forming broad plates similar to a primitive turtle shell, and the vertebrae were nearly identical to those of some turtles. Accordingly, it is often considered as a possible transitional fossil between turtles and their prehistoric ancestors. However, it is possible that these turtle-like features evolved independently of the same features in turtles, since other anatomical studies and phylogenetic analyses suggest that Eunotosaurus may instead have been a parareptile, an early-diverging neodiapsid unrelated to turtles, or a synapsid.
Reptiles arose about 320 million years ago during the Carboniferous period. Reptiles, in the traditional sense of the term, are defined as animals that have scales or scutes, lay land-based hard-shelled eggs, and possess ectothermic metabolisms. So defined, the group is paraphyletic, excluding endothermic animals like birds that are descended from early traditionally-defined reptiles. A definition in accordance with phylogenetic nomenclature, which rejects paraphyletic groups, includes birds while excluding mammals and their synapsid ancestors. So defined, Reptilia is identical to Sauropsida.

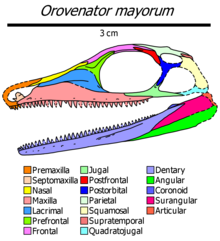
Orovenator is an extinct genus of diapsid from Lower Permian deposits of Oklahoma, United States. It is known from two partial skulls from the Richards Spur locality in Oklahoma. The holotype OMNH 74606 consists of a partial skull preserving snout and mandible, and the referred specimen, OMNH 74607, a partial skull preserving the skull roof, vertebrae and palatal elements. It was first named by Robert R. Reisz, Sean P. Modesto and Diane M. Scott in 2011 and the type species is Orovenator mayorum. The generic name means "mountain", oro, in Greek in reference to the Richards Spur locality, which was mountainous during the Permian period and "hunter", venator, in Latin. The specific name honours Bill and Julie May. Orovenator is the oldest and most basal neodiapsid to date.

Protorosauria is an extinct, likely paraphyletic group of basal archosauromorph reptiles from the latest Middle Permian to the end of the Late Triassic of Asia, Europe and North America. It was named by the English anatomist and paleontologist Thomas Henry Huxley in 1871 as an order, originally to solely contain Protorosaurus. Other names which were once considered equivalent to Protorosauria include Prolacertiformes and Prolacertilia.

Younginidae is an extinct family of diapsid reptiles from the Late Permian and Early Triassic. In a phylogenetic context, younginids are near the base of the clade Neodiapsida. Younginidae includes the species Youngina capensis from the Late Permian of South Africa and Thadeosaurus colcanapi from the Late Permian and Early Triassic of Madagascar. Heleosuchus griesbachi from the Late Permian of South Africa may also be a member of the family.

Pantestudines or Pan-Testudines is the group of all reptiles more closely related to turtles than to any other living animal. It includes both modern turtles and all of their extinct relatives. Pantestudines with a complete shell are placed in the clade Testudinata.

Pappochelys is an extinct genus of diapsid reptile possibly related to turtles. The genus contains only one species, Pappochelys rosinae, from the Middle Triassic of Germany, which was named by paleontologists Rainer Schoch and Hans-Dieter Sues in 2015. The discovery of Pappochelys provides strong support for the placement of turtles within Diapsida, a hypothesis that has long been suggested by molecular data, but never previously by the fossil record. It is morphologically intermediate between the definite stem-turtle Odontochelys from the Late Triassic of China and Eunotosaurus, a reptile from the Middle Permian of South Africa.