An oncogene is a gene that has the potential to cause cancer. In tumor cells, these genes are often mutated, or expressed at high levels.

Angiomotin (AMOT) is a protein that in humans is encoded by the AMOT gene. It belongs to the motin family of angiostatin binding proteins, which includes angiomotin, angiomotin-like 1 (AMOTL1) and angiomotin-like 2 (AMOTL2) characterized by coiled-coil domains at N-terminus and consensus PDZ-binding domain at the C-terminus. Angiomotin is expressed predominantly in endothelial cells of capillaries as well as angiogenic tissues such as placenta and solid tumor.

Growth factor receptor-bound protein 2, also known as Grb2, is an adaptor protein involved in signal transduction/cell communication. In humans, the GRB2 protein is encoded by the GRB2 gene.

Activator protein 1 (AP-1) is a transcription factor that regulates gene expression in response to a variety of stimuli, including cytokines, growth factors, stress, and bacterial and viral infections. AP-1 controls a number of cellular processes including differentiation, proliferation, and apoptosis. The structure of AP-1 is a heterodimer composed of proteins belonging to the c-Fos, c-Jun, ATF and JDP families.
TEAD2, together with TEAD1, defines a novel family of transcription factors, the TEAD family, highly conserved through evolution. TEAD proteins were notably found in Drosophila (Scalloped), C. elegans, S. cerevisiae and A. nidulans. TEAD2 has been less studied than TEAD1 but a few studies revealed its role during development.

Proto-oncogene tyrosine-protein kinase Src, also known as proto-oncogene c-Src, or simply c-Src, is a non-receptor tyrosine kinase protein that in humans is encoded by the SRC gene. It belongs to a family of Src family kinases and is similar to the v-Src gene of Rous sarcoma virus. It includes an SH2 domain, an SH3 domain and a tyrosine kinase domain. Two transcript variants encoding the same protein have been found for this gene.

Proto-oncogene tyrosine-protein kinase Yes is a non-receptor tyrosine kinase that in humans is encoded by the YES1 gene.

Mitogen-activated protein kinase kinase kinase 8 is an enzyme that in humans is encoded by the MAP3K8 gene.

Cell division protein kinase 8 is an enzyme that in humans is encoded by the CDK8 gene.

Transcriptional enhancer factor TEF-1 also known as TEA domain family member 1 (TEAD1) and transcription factor 13 (TCF-13) is a protein that in humans is encoded by the TEAD1 gene. TEAD1 was the first member of the TEAD family of transcription factors to be identified.

Mitogen-activated protein kinase kinase kinase 4 is an enzyme that in humans is encoded by the MAP3K4 gene.
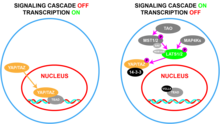
Large tumor suppressor kinase 1 (LATS1) is an enzyme that in humans is encoded by the LATS1 gene.

Serine/threonine-protein kinase 3 is an enzyme that in humans is encoded by the STK3 gene.
Large tumor suppressor kinase 2 (LATS2) is an enzyme that in humans is encoded by the LATS2 gene.

WW domain-containing transcription regulator protein 1 (WWTR1), also known as Transcriptional coactivator with PDZ-binding motif (TAZ), is a protein that in humans is encoded by the WWTR1 gene. WWTR1 acts as a transcriptional coregulator and has no effect on transcription alone. When in complex with transcription factor binding partners, WWTR1 helps promote gene expression in pathways associated with development, cell growth and survival, and inhibiting apoptosis. Aberrant WWTR1 function has been implicated for its role in driving cancers. WWTR1 is often referred to as TAZ due to its initial characterization with the name TAZ. However, WWTR1 (TAZ) is not to be confused with the protein tafazzin, which originally held the official gene symbol TAZ, and is now TAFAZZIN.
The Akt signaling pathway or PI3K-Akt signaling pathway is a signal transduction pathway that promotes survival and growth in response to extracellular signals. Key proteins involved are PI3K and Akt.

The Hippo signaling pathway, also known as the Salvador-Warts-Hippo (SWH) pathway, is a signaling pathway that controls organ size in animals through the regulation of cell proliferation and apoptosis. The pathway takes its name from one of its key signaling components—the protein kinase Hippo (Hpo). Mutations in this gene lead to tissue overgrowth, or a "hippopotamus"-like phenotype.

The WW domain is a modular protein domain that mediates specific interactions with protein ligands. This domain is found in a number of unrelated signaling and structural proteins and may be repeated up to four times in some proteins. Apart from binding preferentially to proteins that are proline-rich, with particular proline-motifs, [AP]-P-P-[AP]-Y, some WW domains bind to phosphoserine- and phosphothreonine-containing motifs.

Marius Sudol is an American molecular and cellular biologist. He was born in 1954 in Tarnow, Poland. In 1978, he immigrated to the United States to study at The Rockefeller University in New York City, where he received his Ph.D. in 1983. He is currently an Adjunct Faulty at the Icahn School of Medicine at Mount Sinai in NYC.
Barry James Thompson is an Australian and British developmental biologist and cancer biologist. Thompson is known for identifying genes, proteins and mechanisms involved in epithelial polarity, morphogenesis and cell signaling via the Wnt and Hippo signaling pathways, which have key roles in human cancer.