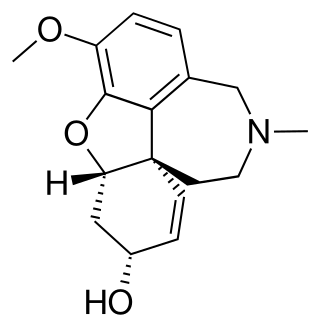
Methyllycaconitine (MLA) is a diterpenoidalkaloid found in many species of Delphinium (larkspurs).[1][2] In common with many other diterpenoid alkaloids, it is toxic to animals, although the acute toxicity varies with species.[3][4] Early research was focused on identifying, and characterizing the properties of methyllycaconitine as one of the principal toxins in larkspurs responsible for livestock poisoning in the mountain rangelands of North America.[3][5] Methyllycaconitine has been explored as a possible therapeutic agent for the treatment of spastic paralysis,[6] and it has been shown to have insecticidal properties.[7] Most recently, it has become an important molecular probe for studying the pharmacology of the nicotinic acetylcholine receptor.[8]
The first isolation of MLA, from Delphinium brownii, Rydb., was probably made by Richard Manske at the National Research Laboratories in Ottawa, Canada, in 1938.[9] Presumably because he did not obtain the compound in sufficiently pure form, Manske declined to give it a name. The name "methyl-lycaconitine" was assigned by John Goodson, working at the Wellcome Chemical Research Laboratories in London, England, when he isolated the alkaloid, in purer form, from seeds of Delphinium elatum, L. in 1943.[10] A more modern isolation procedure is described by Pelletier and his co-workers, who used seeds of the "garden larkspur", Consolida ambigua (also referred to as Delphinium ajacis) as their plant source.[11]
Structure determination
The complete molecular structure for MLA, correct in all but one detail, was first published by Kuzovkov and Platonova in 1959.[12] This structure, supported in part by X-ray crystallography (considered usually to be a "definitive" analytical technique) of a chemical derivative of MLA performed by Maria Przybylska,[13] was accepted as correct until the early 1980s. At that time, the research groups of Pelletier[14] and of Edwards and Przybylska[15] independently corrected the stereochemistry of the methoxy group at C-1 from the β- to α- configuration. Thus any drawing of MLA appearing before Pelletier's 1981 paper[14] will show the structure with the incorrect stereochemistry at C-1.
Chemistry
Synonyms
[1α,4(S),6β,14α,16β]-20-Ethyl-1,6,14,16-tetramethoxy-4-[[[2-(3-methyl-2,5-dioxo-1-pyrrolidinyl)benzoyl]oxy]methyl]aconitane-7,8-diol; also referred to, incorrectly, as "N-methyl lycaconitine" in a few publications.
Physico-chemical properties
MLA is soluble in chloroform, but does not dissolve well in water.[10] The free base of MLA has not been obtained in crystalline form, and in its amorphous form it melts ultimately at 128°C;[10] the hydriodide salt has a melting point of 201°C.;[10] the perchlorate salt melts at 195°C[16] The citrate salt is the most common form in which MLA is currently available commercially.[17]
A pKa does not seem to have been recorded for MLA, but it is considered to be a weak base because it can be readily extracted into diethyl ether from an aqueous solution at pH 7.5-8.[14]
The optical rotation of the free base, [α]D was found to be +49° in alcohol.[10]
Molecular structure
Although commonly referred to as a "diterpenoid" alkaloid, MLA is, strictly speaking, a nor-diterpenoid, since its carbon skeleton only contains 19 C atoms, one having been deleted somewhere during its biosynthesis.[18] Otherwise, the MLA molecule comprises a tertiary amine, two tertiary alcohols, four methylether groups, and a complex ester based on anthranilic acid and methylsuccinic acid. This N-(2-carboxyphenyl)-methylsuccinamido-ester is quite rare amongst natural products.
Synthesis
As of April, 2012 no total synthesis of MLA has been reported. A semi-synthesis of MLA, starting from its "parent" amino-alcohol, lycoctonine (obtained by simple alkaline hydrolysis of natural MLA [10]) was reported in 1994.[19]
Pharmacology
In many respects, the pharmacology of MLA closely resembles that of the classical neuromuscular blocker, d-tubocurarine. The "curare-like" properties of MLA seem to have been first mentioned in 1958 by Kuzovkov and Bocharnikova,[20] working at the Ordzhinikidze All-Union Institute for Scientific Research in Pharmaceutical Chemistry, in the former USSR. A detailed paper on the pharmacology of MLA (in the form of its hydriodide salt, given the drug name "mellictine") in classical animal preparations was published from the same Institute in the following year by Dozortseva.[21]
These studies, together with related others and some original observations, are summarized in the review by Benn and Jacyno.[3]
In the rat phrenic nerve-diaphragm preparation, for example, a 2 x 10−5M concentration of MLA produced a 50% decrease in response, and total inhibition was caused by a 3 x 10−5M concentration of the drug. In this preparation, MLA-treated muscle responded normally to direct electrical stimulation, but the inhibition of contractions was only partially antagonized by physostigmine. Similar results were obtained with frog nerve-muscle preparations, in which it was shown that MLA blocked response of the gastrocnemius muscle to electrical stimulation of the sciatic nerve, inhibited post-synaptic action potentials in the sartorius muscle elicited by stimulation of the sciatic nerve, and reduced the amplitude of miniature end-plate potentials in the extensor digitus IV muscle.
Ganglion-blocking effects of MLA were observed using the cat nictitating membrane preparation: complete inhibition of the response was produced by 4mg/kg of "mellictine" given intravenously.
No significant effects were produced by the drug in smooth muscle preparations from rabbit, guinea pig or cat, indicating the lack of activity at typically muscarinic sites. In electrically stimulated guinea pig ileum, for example, contractions were unaffected by a concentration of 5 x 10−4M of MLA.
A more detailed summary of the above data, together with much related material, may be found in a review written by Kip Panter and collaborators at USDA-ARS laboratories in Utah and California.[22]
A significant advance was made towards understanding the pharmacology of MLA when Jennings and co-workers[7] at the American Cyanamid Company reported that MLA (as its citrate salt) strongly inhibited the binding of tritiated propionyl-α-bungarotoxin to a receptor preparation from house-fly heads, with a Ki of ~ 2.5 x 10−10M. Subsequently, Macallan and his co-workers[23] showed that MLA also competed with 125I-α-bungarotoxin (Ki ~1 x 10−9M) and tritiated (−)-nicotine (Ki ~4 x 10−6M) in a receptor preparation from rat brain. These workers also reported that MLA displaced125I-α-bungarotoxin from purified Torpedo (electric ray) nicotinic acetylcholine receptors (nAChRs) with a Ki ~1 x 10−6M. Similar experiments performed later by Ward et al.[24] showed that MLA bound to nAChRs extracted from human muscle with a Ki of ~8 x 10−6M; it was also reported that MLA, at a concentration of 10−4M, had no affinity for muscarinic AChRs, as labeled by tritiatedquinuclidinyl benzilate, from rat brain.
Further details about the binding of MLA to nAChRs were presented by Wonnacott and her co-workers,[8] who provided evidence that MLA bound preferentially to different sub-units, as expressed in Xenopus frog oocytes, of the nAChR cloned from avian DNA: MLA was found to have an IC50 of ~8 x 10−8M at α3β2 and ~7 x 10−7M at α4β2 receptor sub-types. Although it was also established that MLA bound strongly to α7 sub-types, experimental difficulties precluded the determination of an IC50. Subsequently, research groups from Abbott Laboratories in the USA, and the University of Geneva in Switzerland reported that MLA displaced 125I-α-bungarotoxin from α7 receptors cloned from the human K28 cell line, with a Ki of ~ 1 x 10−8.[25]
One last milestone in the ongoing saga of MLA pharmacology (there are, as of April 2012, approximately 660 references to articles in journals covered by PubMed) to be mentioned is the characterization of the receptor-interactions of tritium-labeled MLA, by researchers at the University of Bath, in the UK.[26]
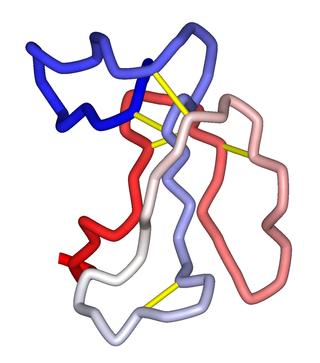
One relatively recent study which sheds light on the interaction of MLA with acetylcholine-binding proteins (AChBP) at the molecular level is that of Hansen et al.,[27] who made observations on the crystal structure of a complex between MLA and an AChBP isolated from the salt-water snail, Aplysia californica.
Toxicology
The toxicology of MLA has been studied largely in the context of livestock poisoning by wild larkspurs. The seminal work by John Jacyno and Mike Benn at the University of Calgary in Canada showed that MLA was most likely to be the agent responsible for the toxicity of a local larkspur, D. brownii, and provided some preliminary acute toxicity data in several animal species.[3][4][5] These LD50s are as follows: mouse, 3–5mg/kg; frog, 3–4mg/kg; rabbit, 2–3mg/kg (after parenteral administration). Cats appeared to have comparable susceptibility to rabbits, whereas dogs were ~ 1.5 x more sensitive.[21] These early observations have been comprehensively extended by USDA researchers,[22] who have estimated the LD50 of MLA to be ~10mg/kg in sheep, ~ 5mg/kg in rats, and ~2mg/kg in cattle.
Although most LD50s are usually determined from parenteral administration of the test drug, MLA is also active when taken orally.[21]
Signs of toxicity in calves, sheep, rats and mice, at low doses, included agitation, respiratory difficulty, and loss of motor control; symptoms appeared within 2–3 minutes of injection, and disappeared within 10 minutes. Doses large enough to produce collapse also caused an increase in heart and respiration rates, as well as tremor, with significant convulsions evident in mice and rats, but not in cattle or sheep.[22] In cases where death seemed imminent, the poisoning in sheep could be counteracted by the i.v. administration of neostigmine and atropine,[22] whereas poisoning in calves was reversed by the administration of physostigmine.[4] In animals that were allowed to die, death appeared to be the result of complete motor paralysis and respiratory arrest.[21][22]
It is worth noting that although a LD50 for man is not available, the clinical studies of Kabelyanskaya showed that an oral dose of 0.02 g of MLA hydriodide ("mellictine") might be given to patients up to 5 times per day, over the course of 1 month. However, some subjects could only tolerate single doses of 0.02 g per day without experiencing side-effects.[6]
Structure-Activity relationships
The earliest observation on a relationship between the molecular structure of MLA and a biological activity concerned the effect of the C-18 ester group on acute toxicity. When this group was hydrolyzed, the resulting amino-alcohol (named lycoctonine as a consequence of its natural occurrence) was found to be much less poisonous to animals than was MLA.[3] A recent study comparing the LD50 of MLA and lycoctonine, given i.v. to mice, showed that lycoctonine was more than 100x less toxic than MLA.[22] In other functional pharmacological assays, lycoctonine resembled MLA qualitatively but was roughly ten times less potent.[3]
When compared in nAChR-binding studies, MLA was found to compete for 125I-α-bungarotoxin binding sites (i.e. α7 sub-types) over 1000x more strongly than did lycoctonine.[28]
If the succinimide ring is deleted so as to leave only the -NH2 group attached to the benzene ring (as in the alkaloid anthranoyllycoctonine, which also occurs naturally), the resulting compound is intermediate between MLA and lycoctonine in potency and toxicity: it is less acutely toxic than MLA by a factor of about 4, but its affinity for 125I-α-bungarotoxin binding sites is over 200x lower than that of MLA.[29]
If the -NH2 group of anthranoyllycoctonine is removed, giving the compound lycoctonine-18-O-benzoate, the affinity for α7 receptors, as well as for α4β2 receptors is reduced by about a factor of 10 in comparison to MLA.[30] When compared with MLA in the rat phrenic nerve-diaphragm assay, lycoctonine-18-O-benzoate was also about 10x less potent, and a similar reduction in potency was observed in an electrophysiological study involving frog extensor muscle.[3]
Even the absence of the methyl group from the methylsuccinimido- ring, as in the alkaloid lycaconitine, reduces the affinity for α7 receptors by a factor of about 20,[31]> but in this case affinity for α4β2 receptors is not significantly changed in comparison with MLA.[30]
Another approach that has been explored in the attempt to elucidate structure-activity relationships in MLA has been to start with 2-(methylsuccinimido)-benzoic acid (the carboxylic acid produced when MLA is split at the C-18 ester group) and to esterify it with various alcohols and amino-alcohols that might be considered as "molecular fragments" of MLA. None of these compounds showed any significant degree of the biological actions characteristic of MLA, however, in the limited number of assays to which they were subjected.[3][22]
Therapeutic applications
MLA has been used for treating a variety of neurological disorders,[6][32] although there are no references to such use in the last few decades.
More recently, it has been proposed that MLA might be useful in reducing nicotine reward without precipitating symptoms of nicotine withdrawal.[33] This suggestion was made on the basis of experiments in which intraperitoneal doses of ~4mg/kg and 8mg/kg of MLA significantly reduced nicotine self-administration in rats.
Most recently, it has been suggested[34] that MLA had potential in the treatment of cannabis dependence. However, this suggestion was apparently based only on work by Solinas et al.[35] who showed that doses of 0.3-5.6mg/kg, i.p., in rats, dose-dependently antagonized the discriminative-stimulus effects of 3mg/kg THC.
Given that the early Soviet work[6] with "mellictine" indicated that as little as ~0.2-0.3mg/kg, orally, in man (assuming a weight of 60–70kg, for the sake of making the dose conversion) could produce symptoms of toxicity, and that oral administration of most drugs typically requires more drug than parenteral administration, it is uncertain if MLA will prove to be a practical treatment for either nicotine or cannabis addiction, based on the effective doses required in the rat experiments.
In a recent review, Wu and co-workers[36] have cited research in which α7-antagonists such as MLA show potential in cancer treatment, but this work is still in its very early stages.
Insecticidal action
Jennings and co-workers, in addition to making their key observations (see Pharmacology above) about the receptor-binding of MLA, found it to be toxic (50+% mortality) to the following insect species: Empoasca abrupta[37] (at 100 ppm), Heliothis virescens (at 1000 ppm), Musca domestica (at 1000 ppm) and Spodoptera eridana (at 1000 ppm). Species which were not significantly affected by MLA were: Anopheles quadrimaculatus, Aphis fabae, Diabrotica undecimpunctuata howardi and Tetranychus urticae. MLA also behaved as a feeding deterrent, with an LC50 of ~300 ppm, to Spodoptera larvae feeding on bean leaves.[7]
Related Research Articles
Aconitine is an alkaloid toxin produced by various plant species belonging to the genus Aconitum, known also commonly by the names wolfsbane and monkshood. Monkshood is notorious for its toxic properties.
A neuromuscular junction is a chemical synapse between a motor neuron and a muscle fiber.
Nicotinic acetylcholine receptors, or nAChRs, are receptor polypeptides that respond to the neurotransmitter acetylcholine. Nicotinic receptors also respond to drugs such as the agonist nicotine. They are found in the central and peripheral nervous system, muscle, and many other tissues of many organisms. At the neuromuscular junction they are the primary receptor in muscle for motor nerve-muscle communication that controls muscle contraction. In the peripheral nervous system: (1) they transmit outgoing signals from the presynaptic to the postsynaptic cells within the sympathetic and parasympathetic nervous system, and (2) they are the receptors found on skeletal muscle that receive acetylcholine released to signal for muscular contraction. In the immune system, nAChRs regulate inflammatory processes and signal through distinct intracellular pathways. In insects, the cholinergic system is limited to the central nervous system.
Galantamine is used for the treatment of cognitive decline in mild to moderate Alzheimer's disease and various other memory impairments. It is an alkaloid that has been isolated from the bulbs and flowers of Galanthus nivalis, Galanthus caucasicus, Galanthus woronowii, and some other members of the family Amaryllidaceae, such as Narcissus (daffodil), Leucojum aestivum (snowflake), and Lycoris including Lycoris radiata. It can also be produced synthetically.
Epibatidine is a chlorinated alkaloid that is secreted by the Ecuadoran frog Epipedobates anthonyi and poison dart frogs from the Ameerega genus. It was discovered by John W. Daly in 1974, but its structure was not fully elucidated until 1992. Whether epibatidine is the first observed example of a chlorinated alkaloid remains controversial, due to challenges in conclusively identifying the compound from the limited samples collected by Daly. By the time that high-resolution spectrometry was used in 1991, there remained less than one milligram of extract from Daly's samples, raising concerns about possible contamination. Samples from other batches of the same species of frog failed to yield epibatidine.
Tubocurarine is a toxic alkaloid historically known for its use as an arrow poison. In the mid-1900s, it was used in conjunction with an anesthetic to provide skeletal muscle relaxation during surgery or mechanical ventilation. Safer alternatives, such as cisatracurium and rocuronium, have largely replaced it as an adjunct for clinical anesthesia and it is now rarely used.
Neuromuscular-blocking drugs, or Neuromuscular blocking agents (NMBAs), block transmission at the neuromuscular junction, causing paralysis of the affected skeletal muscles. This is accomplished via their action on the post-synaptic acetylcholine (Nm) receptors.
α-Bungarotoxin is one of the bungarotoxins, components of the venom of the elapid Taiwanese banded krait snake. It is a type of α-neurotoxin, a neurotoxic protein that is known to bind competitively and in a relatively irreversible manner to the nicotinic acetylcholine receptor found at the neuromuscular junction, causing paralysis, respiratory failure, and death in the victim. It has also been shown to play an antagonistic role in the binding of the α7 nicotinic acetylcholine receptor in the brain, and as such has numerous applications in neuroscience research.
Delphinium nudicaule, known by the common names canyon larkspur, red larkspur, orange larkspur, and canyon delphinium, is a flowering herbaceous perennial plant in the buttercup family Ranunculaceae. It is native to low-elevation canyons and slopes, foothills, and mountain ranges of California, US, from the Sierra Nevada to the California Coast Ranges, and of Oregon. It grows below 6,500 feet (2,000 m).
A nicotinic agonist is a drug that mimics the action of acetylcholine (ACh) at nicotinic acetylcholine receptors (nAChRs). The nAChR is named for its affinity for nicotine.
Bungarotoxins are toxins found in the venom of snakes and kraits. Bites from these animals can result in severe symptoms including bleeding or hemorrhage, paralysis and tissue damage that can result in amputation. The paralytic effects of venom are particularly dangerous as they can impair breathing. These symptoms are the result of bungarotoxin presence in the venom. In actuality, venom contains several distinct bungarotoxins, each varying in which receptors they act on and how powerful they are.
α-Cobratoxin is a substance of the venom of certain Naja cobras. It is a nicotinic acetylcholine receptor (nAChR) antagonist which causes paralysis by preventing the binding of acetylcholine to the nAChR.
Surugatoxin (SGTX) is a type of venom found in the mid-gut digestive gland of the Japanese ivory mollusk Babyloniajaponica, a carnivorous gastropod. It functions as a ganglionic blocker of nicotinic acetylcholine receptors (nAChRs). The structurally and functionally related neosurugatoxin, also derived from Babylonia japonica, is an even more potent nAChR antagonist than SGTX.
Delphinine is a toxic diterpenoid alkaloid found in plants from the Delphinium (larkspur) and Atragene genera, both in the family Ranunculaceae. Delphinine is the principal alkaloid found in Delphinium staphisagria seeds – at one time, under the name stavesacre, a very well known herbal treatment for body lice. It is related in structure and has similar effects to aconitine, acting as an allosteric modulator of voltage gated sodium channels, and producing low blood pressure, slowed heart rate and abnormal heart rhythms. These effects make it highly poisonous. While it has been used in some alternative medicines, most of the medical community does not recommend using it due to its extreme toxicity.
α-Neurotoxins are a group of neurotoxic peptides found in the venom of snakes in the families Elapidae and Hydrophiidae. They can cause paralysis, respiratory failure, and death. Members of the three-finger toxin protein family, they are antagonists of post-synaptic nicotinic acetylcholine receptors (nAChRs) in the neuromuscular synapse that bind competitively and irreversibly, preventing synaptic acetylcholine (ACh) from opening the ion channel. Over 100 α-neurotoxins have been identified and sequenced.
3-Bromocytisine is a derivative of the toxic alkaloid cytisine that acts as a highly potent agonist at neural nicotinic acetylcholine receptors, binding primarily to the α4β2 and α7 subtypes. 3-Bromocytisine is a full agonist at the α7 subtype while it is only a partial agonist at α4β2, but has an extremely strong binding affinity at α4β2 with 200-fold selectivity for α4β2 over α7. In animal studies 3-bromocytisine stimulates the release of dopamine and noradrenaline and increases locomotor activity.
κ-Bungarotoxin is a protein neurotoxin of the bungarotoxin family that is found in the venom of the many-banded krait, a snake found in Taiwan. κ-Bungarotoxin is a high affinity antagonist of nicotinic acetylcholine receptors (nAChRs), particularly of CHRNA3; it causes a post-synaptic blockade of neurotransmission. Although there is significant variability in the clinical effects of snake bites, neuromuscular paralysis and respiratory failure are associated with krait bites.
Dioscorine is an alkaloid toxin isolated from the tubers of tropical yam on several continents. It has been used as a monkey poison in some African countries, and as an arrow poison to aid in hunting in several parts of Asia. It was first isolated from Dioscorea hirsute by Boorsma in 1894 and obtained in a crystalline form by Schutte in 1897, and has since been found in other Dioscorea species. Dioscorine is a neurotoxin that acts by blocking the nicotinic acetylcholine receptor. Dioscorine is generally isolated in tandem with other alkaloids such as dioscin but is usually the most potent toxin in the mixture. It is a convulsant, producing symptoms similar to picrotoxin, with which it shares a similar mechanism of action.
Three-finger toxins are a protein superfamily of small toxin proteins found in the venom of snakes. Three-finger toxins are in turn members of a larger superfamily of three-finger protein domains which includes non-toxic proteins that share a similar protein fold. The group is named for its common structure consisting of three beta strand loops connected to a central core containing four conserved disulfide bonds. The 3FP protein domain has no enzymatic activity and is typically between 60-74 amino acid residues long. Despite their conserved structure, three-finger toxin proteins have a wide range of pharmacological effects. Most members of the family are neurotoxins that act on cholinergic intercellular signaling; the alpha-neurotoxin family interacts with muscle nicotinic acetylcholine receptors (nAChRs), the kappa-bungarotoxin family with neuronal nAChRs, and muscarinic toxins with muscarinic acetylcholine receptors (mAChRs).
Azemiopsin, a toxin obtained from the Azemiops feae viper venom, is a polypeptide that consists of 21 amino acid residues. It does not contain cysteine residues or disulfide bridges. The polypeptide can block skeletal muscle contraction by blocking nicotinic acetylcholine receptors.
1 2 Aiyar VN, Benn MH, Hanna T, Jacyno J, Roth SH, Wilkens JL (October 1979). "The principal toxin of Delphinium brownii Rydb., and its mode of action". Experientia. 35 (10): 1367–8. doi:10.1007/BF01964013. PMID499426. S2CID19818861.
1 2 3 4 Kabelianskaia LG (1959). "[Use of mellictin in clinical diseases of the nervous system in spastic paralysis]". Farmakologiia I Toksikologiia. 22 (1): 38–42. PMID13653123.
1 2 3 Jennings KR, Brown DG, Wright DP (1986). "Methyllycaconitine, a naturally occurring insecticide with a high affinity for the insect cholinergic receptor". Experientia. 42 (6): 611–613. doi:10.1007/BF01955557. S2CID1460936.
1 2 Wonnacott E, Albuquerque X, Bertrand D (1993). Conn PM (ed.). Methods in Neurosciences. Vol.12. San Diego: Academic Press. pp.263–275.
↑ Manske RH (March 1939). "Missing". Canadian Journal of Research. 16: 57–60. doi:10.1139/cjr39b-010.
1 2 3 4 5 6 Goodson JA (1943). "42. The alkaloids of the seeds of Delphinium elatum, L.". Journal of the Chemical Society (Resumed): 139–41. doi:10.1039/JR9430000139.
↑ Pelletier SW, Sawhney RS, Desai HK, Mody NV (May 1980). "The diterpenoid alkaloids of Consolida ambigua". Journal of Natural Products. 43 (3): 395–406. doi:10.1021/np50009a011.
↑ Kuzovkov AD, Platonova TF (1959). "Missing". J. Gen. Chem. (Eng. Trans.). 29: 2746–2749.
↑ Przybylska M, Marion L (February 1956). "The Crystal Structure of Des-(oxymethylene)_lycoctonine Hydroidodide Monohydrate". Canadian Journal of Chemistry. 34 (2): 185–7. doi:10.1139/v56-026.
1 2 3 Pelletier SW, Mody NV, Varughese KI, Maddry JA, Desai HK (October 1981). "Structure revision of 37 lycoctonine-related diterpenoid alkaloids". Journal of the American Chemical Society. 103 (21): 6536–8. doi:10.1021/ja00411a062.
↑ Gopalakrishnan M, Buisson B, Touma E, Giordano T, Campbell JE, Hu IC, etal. (August 1995). "Stable expression and pharmacological properties of the human alpha 7 nicotinic acetylcholine receptor". European Journal of Pharmacology. 290 (3): 237–46. doi:10.1016/0922-4106(95)00083-6. PMID7589218.
↑ Davies AR, Hardick DJ, Blagbrough IS, Potter BV, Wolstenholme AJ, Wonnacott S (May 1999). "Characterisation of the binding of [3H]methyllycaconitine: a new radioligand for labelling alpha 7-type neuronal nicotinic acetylcholine receptors". Neuropharmacology. 38 (5): 679–90. doi:10.1016/s0028-3908(98)00221-4. PMID10340305. S2CID23349607.
↑ Coates PA, Blagbrough IS, Hardick DJ, Rowan MG, Wonnacott S, Potter BV (November 1994). "Rapid and efficient isolation of the nicotinic receptor antagonist methyllycaconitine from Delphinium: Assignment of the methylsuccinimide absolute stereochemistry as S.". Tetrahedron Letters. 35 (46): 8701–4. doi:10.1016/S0040-4039(00)78476-0.
↑ Hardick DJ, Blagbrough IS, Cooper G, Potter BV, Critchley T, Wonnacott S (November 1996). "Nudicauline and elatine as potent norditerpenoid ligands at rat neuronal alpha-bungarotoxin binding sites: importance of the 2-(methylsuccinimido)benzoyl moiety for neuronal nicotinic acetylcholine receptor binding". Journal of Medicinal Chemistry. 39 (24): 4860–6. doi:10.1021/jm9604991. PMID8941400.
1 2 Jacyno JM, etal. (1995). Gustine DL, Flores HE (eds.). Phytochemicals and Health. Current Topics in Plant Physiology. Vol.15. Rockville: American Society of Plant Physiologists. pp.294–295.
↑ Jacyno JM, Harwood JS, Lin NH, Campbell JE, Sullivan JP, Holladay MW (July 1996). "Lycaconitine revisited: partial synthesis and neuronal nicotinic acetylcholine receptor affinities". Journal of Natural Products. 59 (7): 707–9. doi:10.1021/np960352c. PMID8759171.
↑ Gubanov IA (1965). "Missing". Planta Medica. 13: 200–205.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.