The signal recognition particle (SRP) is an abundant, cytosolic, universally conserved ribonucleoprotein that recognizes and targets specific proteins to the endoplasmic reticulum in eukaryotes and the plasma membrane in prokaryotes.
A signal peptide is a short peptide present at the N-terminus of most newly synthesized proteins that are destined toward the secretory pathway. These proteins include those that reside either inside certain organelles, secreted from the cell, or inserted into most cellular membranes. Although most type I membrane-bound proteins have signal peptides, the majority of type II and multi-spanning membrane-bound proteins are targeted to the secretory pathway by their first transmembrane domain, which biochemically resembles a signal sequence except that it is not cleaved. They are a kind of target peptide.

Cajal bodies (CBs) also coiled bodies, are spherical nuclear bodies of 0.3–1.0 µm in diameter found in the nucleus of proliferative cells like embryonic cells and tumor cells, or metabolically active cells like neurons. CBs are membrane-less organelles and largely consist of proteins and RNA. They were first reported by Santiago Ramón y Cajal in 1903, who called them nucleolar accessory bodies due to their association with the nucleoli in neuronal cells. They were rediscovered with the use of the electron microscope (EM) and named coiled bodies, according to their appearance as coiled threads on EM images, and later renamed after their discoverer. Research on CBs was accelerated after discovery and cloning of the marker protein p80/Coilin. CBs have been implicated in RNA-related metabolic processes such as the biogenesis, maturation and recycling of snRNPs, histone mRNA processing and telomere maintenance. CBs assemble RNA which is used by telomerase to add nucleotides to the ends of telomeres.

Nucleoproteins are proteins conjugated with nucleic acids. Typical nucleoproteins include ribosomes, nucleosomes and viral nucleocapsid proteins.
In cellular biology, P-bodies, or processing bodies, are distinct foci formed by phase separation within the cytoplasm of a eukaryotic cell consisting of many enzymes involved in mRNA turnover. P-bodies are highly conserved structures and have been observed in somatic cells originating from vertebrates and invertebrates, plants and yeast. To date, P-bodies have been demonstrated to play fundamental roles in general mRNA decay, nonsense-mediated mRNA decay, adenylate-uridylate-rich element mediated mRNA decay, and microRNA (miRNA) induced mRNA silencing. Not all mRNAs which enter P-bodies are degraded, as it has been demonstrated that some mRNAs can exit P-bodies and re-initiate translation. Purification and sequencing of the mRNA from purified processing bodies showed that these mRNAs are largely translationally repressed upstream of translation initiation and are protected from 5' mRNA decay.
Heterogeneous nuclear ribonucleoproteins (hnRNPs) are complexes of RNA and protein present in the cell nucleus during gene transcription and subsequent post-transcriptional modification of the newly synthesized RNA (pre-mRNA). The presence of the proteins bound to a pre-mRNA molecule serves as a signal that the pre-mRNA is not yet fully processed and therefore not ready for export to the cytoplasm. Since most mature RNA is exported from the nucleus relatively quickly, most RNA-binding protein in the nucleus exist as heterogeneous ribonucleoprotein particles. After splicing has occurred, the proteins remain bound to spliced introns and target them for degradation.

In molecular biology, LSm proteins are a family of RNA-binding proteins found in virtually every cellular organism. LSm is a contraction of 'like Sm', because the first identified members of the LSm protein family were the Sm proteins. LSm proteins are defined by a characteristic three-dimensional structure and their assembly into rings of six or seven individual LSm protein molecules, and play a large number of various roles in mRNA processing and regulation.
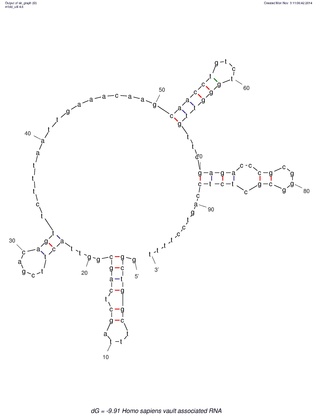
Many eukaryotic cells contain large ribonucleoprotein particles in the cytoplasm known as vaults. The vault complex comprises the major vault protein (MVP), two minor vault proteins, and a variety of small untranslated RNA molecules known as vault RNAs only found in higher eukaryotes. These molecules are transcribed by RNA polymerase III.

snRNP70 also known as U1 small nuclear ribonucleoprotein 70 kDa is a protein that in humans is encoded by the SNRNP70 gene. snRNP70 is a small nuclear ribonucleoprotein that associates with U1 spliceosomal RNA, forming the U1snRNP a core component of the spliceosome. The U1-70K protein and other components of the spliceosome complex form detergent-insoluble aggregates in both sporadic and familial human cases of Alzheimer's disease. U1-70K co-localizes with Tau in neurofibrillary tangles in Alzheimer's disease.

Small nuclear ribonucleoprotein Sm D1 is a protein that in humans is encoded by the SNRPD1 gene.

Major vault protein is a protein that in humans is encoded by the MVP gene. 78 copies of the protein assemble into the large compartments called vaults.

Small nuclear ribonucleoprotein-associated proteins B and B' is a protein that in humans is encoded by the SNRPB gene.

Small nuclear ribonucleoprotein Sm D2 is a protein that in humans is encoded by the SNRPD2 gene. It belongs to the small nuclear ribonucleoprotein core protein family, and is required for pre-mRNA splicing and small nuclear ribonucleoprotein biogenesis. Alternative splicing occurs at this locus and two transcript variants encoding the same protein have been identified.

Small nuclear ribonucleoprotein E is a protein that in humans is encoded by the SNRPE gene.

Small nuclear ribonucleoprotein F is a protein that in humans is encoded by the SNRPF gene.

Poly [ADP-ribose] polymerase 4 is an enzyme that in humans is encoded by the PARP4 gene.

U2 small nuclear ribonucleoprotein B is a protein that in humans is encoded by the SNRPB2 gene.

Telomerase protein component 1 is an enzyme that in humans is encoded by the TEP1 gene.
Leonard H. Rome is a cell biologist and biochemist who has been a faculty member of the David Geffen School of Medicine at UCLA, since he joined the Department of Biological Chemistry there, in 1979. He became a full professor in 1988 and has also served as the Senior Associate Dean for Research in the Geffen School of Medicine from 1997 to 2012. He is the Associate Director of the California NanoSystems Institute (CNSI) since 2004, and was Interim Director from 2007-2009. In addition, he served from 2001 to 2005 as University of California, Los Angeles (UCLA) Associate Vice Chancellor for Research for the Life and Health Sciences.
Nancy Kedersha is an American cell biologist and micrographer. She got her Ph.D. from Rutgers University where she worked in Richard Berg's lab studying the characteristics and assembly of prolyl hydroxylases. Afterwards she joined Leonard Rome's lab at UCLA as a post-doctoral fellow where she co-discovered the vault (organelle). Subsequently, she worked at ImmunoGen Inc. where she worked on staining and photographing different cancer cells. Currently, she works as an instructor of medicine at Brigham and Women's Hospital in Paul Anderson's lab where her work focuses on studying stress granules formation. In addition to her contributions as a scientist, Kedersha has been quite successful in different microscopy competitions. She is a four-time Nikon Small World finalist and in 2011 she won the Lennart Nilsson Award.
