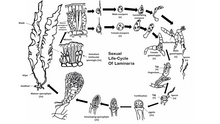
Kelps are large brown algae or seaweeds that make up the order Laminariales. There are about 30 different genera. Despite its appearance, kelp is not a plant but a stramenopile, a group containing many protists.

Nereocystis is a monotypic genus of subtidal kelp containing the species Nereocystis luetkeana. Some English names include edible kelp, bull kelp, bullwhip kelp, ribbon kelp, bladder wrack, and variations of these names. Due to the English name, bull kelp can be confused with southern bull kelps, which are found in the Southern Hemisphere. Nereocystis luetkeana forms thick beds on subtidal rocks, and is an important part of kelp forests.

Laminaria is a genus of brown seaweed in the order Laminariales (kelp), comprising 31 species native to the north Atlantic and northern Pacific Oceans. This economically important genus is characterized by long, leathery laminae and relatively large size. Some species are called Devil's apron, due to their shape, or sea colander, due to the perforations present on the lamina. Others are referred to as tangle. Laminaria form a habitat for many fish and invertebrates.

Macrocystis is a monospecific genus of kelp with all species now synonymous with Macrocystis pyrifera. It is commonly known as giant kelp or bladder kelp. This genus contains the largest of all the Phaeophyceae or brown algae. Macrocystis has pneumatocysts at the base of its blades. Sporophytes are perennial and the individual may live for up to three years; stipes/fronds within a whole individual undergo senescence, where each frond may persist for approximately 100 days. The genus is found widely in subtropical, temperate, and sub-Antarctic oceans of the Southern Hemisphere and in the northeast Pacific from Baja California to Sitka, Alaska. Macrocystis is often a major component of temperate kelp forests.

The Fucales (fucoids) are an order in the brown algae. The list of families in the Fucales, as well as additional taxonomic information on algae, is publicly accessible at Algaebase.

Sporochnacaeae is the only family in the order Sporochnales in the brown algae. Member of this family are thread-like algae growing by means of an intercalary row of dome shaped cells at the base of the hairs.

Ectocarpales is a very large order in the brown algae. The order includes families with pseudoparenchymatous (Splachnidiaceae) or true parenchymatous (Scytosiphonaceae) tissue. Pseudoparenchymatous refers to a filamentous alga with cells packed very close together to give an appearance of parenchymatous tissue, the latter being composed of cells which can truly divide in three dimensions, unusual among the algae. Filamentous algae are composed of cells that divide along a single plane, allowing only elongation to form filaments of one or more rows of cells. Algae that can divide in two planes can form sheet-like thalli or bodies. Cells that can divide in a third plane potentially allow for the organism to develop a more complex body plan, and diversification of body plans into an erect thallus of some sort and a holdfast for attaching the upright portion to the substrate.

Postelsia palmaeformis, also known as the sea palm or palm seaweed, is a species of kelp and classified within brown algae. It is the only known species in the genus Postelsia. The sea palm is found along the western coast of North America, on rocky shores with constant waves. It is one of the few algae that can survive and remain erect out of the water; in fact, it spends most of its life cycle exposed to the air. It is an annual, and edible, though harvesting of the alga is discouraged due to the species' sensitivity to overharvesting.

Sargassaceae is a family of brown algae in the order Fucales.

Eisenia arborea, or the southern sea palm, is a dominant species of kelp that is found on the western Pacific coast of North America, from Vancouver Island, Canada south to Mexico's Isla Magdalena and Baja California, as well as in Japan. They are commonly found from the midtidal areas stretching to the subtidal areas. It is an edible seaweed, a source of nutrients for grazing marine invertebrates and a source of alginic acid, a food thickener. Some of the algas have a hollow stipe above its holdfast with two branches terminating in multiple blades. Eisenia arborea is studied in order to predict environmental stress in oceans intertidal zones. Hollow stipes where present when the Eisenia arborea did not receive essential nutrients for its thalli development. Eisenia arborea with hollow stripes are believed to be evolved algae in order to increase their survival in harsh living conditions. They play a huge role in determining environmental stress.

Durvillaea antarctica, also known as cochayuyo and rimurapa, is a large, robust species of southern bull kelp found on the coasts of Chile, southern New Zealand, and Macquarie Island. D. antarctica, an alga, does not have air bladders, but floats due to a unique honeycomb structure within the alga's blades, which also helps the kelp avoid being damaged by the strong waves.

Egregia menziesii is a species of kelp known commonly as feather boa kelp. It is the only species in the monotypic genus Egregia. It is native to the coastline of western North America from Alaska to Baja California, where it is a common kelp of the intertidal zone.

Choristocarpaceae is a family in the order Discosporangiales of the brown algae. The family contains a single genus, Choristocarpus. The species is mostly located in the cold waters of the Northern hemisphere. A type of seaweed, Choristocarpaceae attaches itself to rocky substrate in places that are near continental shelves and the shore. Due to the species having morphological similarity, they were classified in closer relation with D. mesarthrocarpum. But due to many other differing characteristics Choristocarpaceae were put into their own family with a single genus and a single species of brown algae.
Pterygophora californica is a large species of kelp, commonly known as stalked kelp. It is the only species in its genus Pterygophora. It grows in shallow water on the Pacific coast of North America where it forms part of a biodiverse community in a "kelp forest". It is sometimes also referred to as woody-stemmed kelp, walking kelp, or winged kelp.

Himanthalia is a genus of brown algae. It is the only genus in the family Himanthaliaceae in the order Fucales. It includes two species: Himanthalia durvillei and Himanthalia elongata.
Sunamphitoe femorata is a species of amphipod crustacean in the family Ampithoidae. It is a herbivore and constructs a tubular nest-like home on a blade of the sporophyte of the giant kelp Macrocystis pyrifera. This home is made by rolling the sides of the blade together and securing them with silk. As the kelp blade grows, the nest is advanced down the blade towards the base, approximately keeping pace with the algal growth.
Pleurophycus gardneri is a species of brown alga. It is a deciduous kelp, primarily found in lower, rocky inter-tidal and shallow, rocky sub-tidal locations and is one of the most abundant kelps found within the Pleurophycus Zone. It is not commonly present deeper in the ocean than 30m and is considered a stipitate kelp. P. gardneri forms aggregates of densities up to 10m−2. These kelp beds reside below giant kelp forests, and were therefore often overlooked by researchers for many years. This kelp has a range from Central California to British Columbia, Canada, with a lifespan of only 3 – 6 years.
Fucophycidae is a subclass of Phaeophyceae which contains the most complex and evolved orders of Chromista algae. The members of this subclass have stalks with several morphological forms and distinct structures, characterized by an intercalary growth and a basic heteromorphic, sometimes secondarily iso- or sub-isomorphic life cycle.

Padina is a genus of brown macroalgae in the family Dictyotaceae.