Chromera velia, also known as a "chromerid",[2][1] is a unicellular photosynthetic organism[3] in the superphylum Alveolata. It is of interest in the study of apicomplexan parasites, specifically their evolution and accordingly, their unique vulnerabilities to drugs.[4]
The discovery of C. velia has sparked renewed interest in protist research, concerning both algae and parasites, as well as free-living unicells. Strict separation of botanical protists (algae) and zoological protists (protozoa) has been conventional but C. velia may be regarded as a good example of a bridge linking both categories.[2]
C. velia has typical features of alveolates, being phylogenetically related to Apicomplexa (a subgroup of alveolates), and contains a photosynthetic plastid (chloroplast) while the apicomplexans have a non-photosynthetic plastid called the apicoplast. C. velia is also related to another subgroup of alveolates, the dinoflagellates of which most are photosynthetic.[2]
C. velia uses metabolites (reduced carbon) from its plastid as its primary energy source. The same is true of the algal cousin of C. velia, another chromerid Vitrella brassicaformis.[5] Together these are phylogenetically the closest known autotrophic organisms to apicomplexans.[2][5][6]
Parasites in the apicomplexan genus Plasmodium are the causative agents of malaria. Studies of C. velia and V. brassicaformis are broadly useful for understanding the biochemistry, physiology and evolution of the malaria parasite, other apicomplexan parasites, and dinoflagellates.[2]
Plastid terminology
"Apicoplast" is a specialised word, derived from the word "plastid". Initially the word plastid was more suitable than "chloroplast" when describing organelles of apparent algal descent in any protist, but that lack any chlorophyll or light absorbing pigment. Those found in apicomplexan parasites are a prominent example. The majority of members of the apicomplexan lineage still contain a genome in the plastid, indicating the organelle of the lineage's ancestors was once photosynthetic,[7][8] but these plastids have no light absorbing pigments or light reaction machinery.[8]
While Chromera velia contains a photosynthetic plastid, the majority of apicomplexan relatives contain a non-photosynthetic plastid, and the remainder contain no plastid. The ancestral photosynthetic plastid of ancestral apicomplexans may have been very similar to the plastid of C. velia or the plastid of V. brassicaformis.[6]
Just as the term "plastid" has become widely adopted for chloroplast-derived organelles of non-photosynthetic protists, the term "apicoplast" has also gained acceptance for the plastid of apicomplexans. In current usage, the term plastid may even be used to describe the chloroplast of any photosynthetic organism, and so has a general non-discriminatory use.[6]
With the use of DNA sequencing, a relationship between C. velia, dinoflagellates and apicomplexans was noted.[2] Genomic DNA of C. velia was extracted to provide PCR templates, and when the sequences of the amplified genes were compared with those of other species, biostatistical methods resulted in placement of C. velia on a phylogenetic branch close to the apicomplexans.[2] Through a variety of phylogenetic tests on the orthologous genes found in similar organisms, researchers were able to relate C. velia to dinoflagellates and apicomplexans which are alveolates. Both the nucleus and the plastid of C. velia showed alveolate ancestry. A subsequent study of the C.velia and V. brassicaformis plastid genomes has shown in greater detail that the plastids of peridinin dinoflagellates, apicomplexans and chromerids share the same lineage, derived from a red-algal-type plastid.[6]
Description and availability
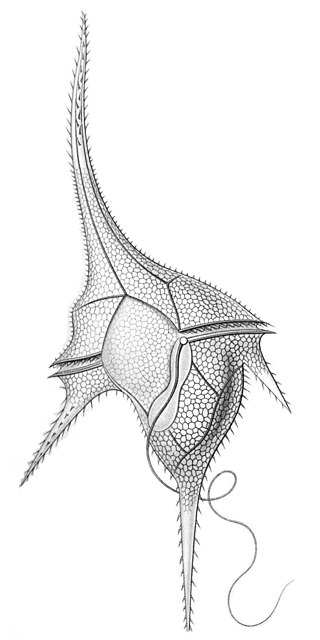
After the naming of the organism and description of the immotile form,[2] several papers have since documented the vegetative motile form[1][9] which excysts in a set of eight siblings from the progenitor cell.[10]
A structure resembling an apical complex in the flagellate,[11] includes a conoid or pseudoconoid and long sacculate micronemes,[1][11] confirming a relationship to apicomplexans. However, this relationship has yet to be formalised, beyond the fact that chromerids and apicomplexans are classified as sister groups within the Alveolata.[2] The precise function of the apical organelles of the Chromerida, is unknown though the organelles have been studied in some detail.[11][12]
Live C. velia is available to purchase from the NCMA culture collection in Maine USA,[13] and is backed up in other culture collections such as CCAP (UK),[14] and SCCAP (Scandinavia).[15]
Preserved material is deposited in the Australian Museum, Sydney, as holotype/hapantotype Z.6967, being a preserved culture embedded in PolyBed 812,[2] and is separately deposited also in absolute ethanol.[2]
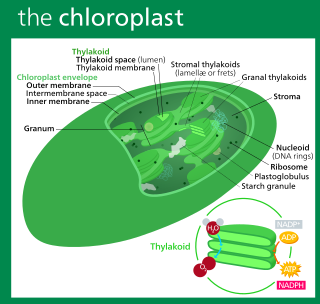
Special features of the C. velia plastid
The plastid of Chromera velia has 4 surrounding membranes and contains chlorophyll a, while chlorophyll c is missing. Photosynthesis has been examined in C. velia, and its photosynthetic carbon assimilation was shown to be very efficient, in the sense of adaptability to a wide range of light regimes, from high light to low light.[16] Thus like other algae that contain only chlorophyll a (such as Nannochloropsis, a stramenopile), the lack of chlorophyll c does not appear to debilitate chromerids in any way. Accessory pigments in C. velia include isofucoxanthin.[2]
Unlike other eukaryotic algae which use only UGG codons to encode the amino acid tryptophan in plastid genomes, the plastid genome of C. velia contains the codon UGA at several positions that encode tryptophan in the psbA gene[2] and other genes.[6] The UGA-Trp codon is characteristic of apicoplasts, and the mitochondria of various organisms, but until the discovery of C. velia, was unprecedented in any photosynthetic plastid. Similarly a bias towards poly-U tails is found specifically on the subset of apicoplast-encoded genes that are involved in photosynthesis in C. velia.[17] Discovery of these two genetic features, the UGA-Trp, and the poly-U tailed photosynthesis genes, indicates that C. velia provides an appropriate model to study the evolution of the apicoplast. Another characteristic feature of C. velia is that its plastid genome is linear-mapping.[18] Janouškovec et al 2013 also presents the expression pathway DNA → RNA →photosystem I protein A1. It is unusually late to fully resolve: It is not fully assembled as a single transcript or even as a single translation product, but only after that step.[19]
Mitochondrion
The mitochondrial genome of C. velia encodes a single gene - cox1 - and several fragmented rRNA molecules.[20][21][22] This mitochondrial genome is one step further devolved than those of peridinin dinoflagellates, which contain three protein-coding genes. However both lineages, C. velia and dinoflagellates, contain functioning mitochondria, the genes having moved to the nucleus.[citation needed]
Most of the Apicomplexan mitochondria that have been previously sequenced also have only three protein encoding genes including cox1 and a number of fragmented rRNA genes. Exceptions to this rule are known: the apicomplexan organism Cryptosporidium appears to lack a mitochondrion entirely.[citation needed]
The C. velia mitochondrial apparatus differs significantly from that of the other chromerid Vitrella brassicaformis. A recent finding is that the respiratory complexes I and III of C. velia are missing, and that the function of complex III has been taken over by a lactate->cytochrome C oxidoreductase [22] By contrast the more ancestral chromerid mitochondrial genome, represented by that of V. brassicaformis retains a canonical complex III.[23]
An unexpected finding in Chromera was a large (1 μm diameter) ever-present organelle bounded by two membranes, originally thought to be the mitochondrion.[2] This organelle may not be a mitochondrion, but an extrusosome called the "chromerosome".[1] The actual mitochondria, by contrast, were found to be small and multiple,[1] just as for other alveolates.[citation needed]
Evolution
The discovery of Chromera velia and its unique plastid which is similar in origin to the apicoplasts, provides an important link in the evolutionary history of the apicomplexans. Previous to the description of C. velia, much speculation surrounded the idea of a photosynthetic ancestral lineage for apicomplexan parasites. For a step by step history of the characterization of the apicomplexan apicoplast organelle, see for example the web review by Vargas Parada (2010).[8]
It is hypothesized that apicomplexans, with their relic chloroplast, the apicoplast, were once able to synthesize energy via photosynthesis. Ancient apicomplexans or their immediate progenitors may have had a symbiotic relationship with the coral reef around them. To achieve that, these ancient organisms would have possessed a working chloroplast. However, if so, this autotrophic ability was lost and apicomplexans have slowly evolved to become parasitic species dependent on their hosts for survival.[24]
Although researchers are still discussing why apicomplexans would sacrifice their photosynthetic ability and become parasitic, it is suggested that clues might be gathered by studying aspects of the evolution of the Chromerida, such as the development of an apical complex of organelles that were used by later descendants to invade host cells.[11][12] In July 2015 the full genome sequences of chromerids C.velia and V. brassicaformis were published,[25] revealing the array of genes that were co-opted or adapted[26] in the transition from a free living lifestyle to a parasitic lifestyle.[26]
The plastid genome of C. velia is unusual in that there is evidence it may be linear [18] and contains split genes for key photosystem genes. The linear state of the C. velia plastid genome is a reminder that C. velia is not an ancestral organism, but is a derived form, which evolved from an ancestral photosynthetic alveolate that presumably had a circular plastid genome, just as the other known chromerid Vitrella brassicaformis does.[citation needed]
Much research surrounds the flagellar apparatus of Chromera, Vitrella and apicomplexans, in relation to the morphological transition of this organelle during the origination of parasitism in apicomplexans.[12][27] It does appear that C. velia exist as a free-living phototroph when necessary or when environmental conditions are suitable, but can also infect coral larvae and live as an intracellular parasite.[28][29]
Pharmacological significance
One potentially important contribution of research on C. velia, besides its position as a missing link between parasitic and algal species, is its potential in studies aimed at finding new antimalarial drugs or clarifying the function of existing antimalarial drugs . Many drugs that have been in clinical use for a long time affect functions in the apicoplast in Plasmodium cells.[30][31] The essential biological function of the apicoplast is solely the production of isoprenoids and their derivatives,[32] without which the parasites cannot live.[32]
C. velia could serve as a convenient model target for the development of antimalarial drugs, since it effectively contains the original apicoplast, as it were, and since its nuclear genome closely resembles that of the ancestral proto-parasites. In the laboratory setting, working with apicomplexan parasites can be difficult, hazardous and expensive, because they must be infected into live host cells (in tissue culture) to remain viable. Chromera velia, is more easily maintained than apicomplexan parasites, yet is related to them, so may potentially provide a laboratory model for the understanding or development of antimalarial treatments. C. velia is able to live independently of its normal animal hosts and can be grown easily and cheaply in a laboratory setting.[citation needed]
Just as humans are subject to infections by the apicomplexans Plasmodium and Cryptosporidium, animals are also subject to infection by apicomplexans including Toxoplasma, Babesia, Neospora, and Eimeria. It is said anecdotally, that almost every animal on earth has one or more species of apicomplexan parasite that challenge it. The economic burden from apicomplexan parasites is estimated in the billions of dollars,[33][34] (see also Malaria) on top of the human and animal costs of these organisms. An increased understanding of the evolutionary roles and functions of apicoplasts and apical complexes can impact on research about the apicomplexan parasites of livestock animals, making C. velia of interest in an agricultural context as well as in the medical and ecological fields.[citation needed]
A provisional patent on the use of Chromerida (Chromera and Vitrella) as subjects for screening and testing of anti-apicomplexan drugs[4] was not lodged as a full patent, which leaves the way open for use of these organisms in commercial development of screening methods for useful compounds.[citation needed]
Ecology
One study has shown that Chromera may have a symbiotic role within corals, being vertically transmitted from parent to offspring Montiporadigitata via the coral's egg stage.[35] The Chromera cells could be cultured from the M.digitata eggs and were subsequently used to transiently colonise Acropora coral larvae. Chromera's known host range therefore includes the corals M. digitata, P. versipora (type host) and L. purpurea (alternate host), and extends through tropical and temperate waters.[2] The symbiont may obtain metabolites from the host,[36] and it has been proposed this may potentially increase its growth rate inside the host.[citation needed]
Analysis of environmental metagenomic datasets has revealed that there are other species related to C. velia and V. brassicaformis associated with corals, but yet to be described.[37] These associations are globally distributed.[37] Among these is the uncultured undescribed "apicomplexan-related lineage-V"[38] which was inferred by the authors to be potentially photosynthetic, and appears to be a symbiosis specialist. Cultured chromerids by comparison can be hypothesized to move between the free-living and coral-associated states, as they are found in M. digitata eggs[35] but are also associated with seaweed, judging from correlations in macroalgal metagenomic datasets.[38] The range of life strategies and niches adopted by apicomplexan-related algae therefore resembles the spectrum of niches occupied by the coral symbiont Symbiodinium.[citation needed]
Research Community
The first Chromera conference and workshop was held at the Heron Island Research Station, Queensland, Australia from November 21–25, 2011.[39] Highlights included diving and culturing. Presentations included the announcement of a formal description of the second isolated chromerid, Vitrella brassicaformis. Professors and students alike participated in the conference and workshop, and a broad range of topics was covered. It was agreed that further meetings would follow. The second conference was held in South Bohemia, Czech Republic, from June 22–25, 2014, arranged by the Oborník lab, via open email list.[citation needed]
Related Research Articles
The Apicomplexa are organisms of a large phylum of mainly parasitic alveolates. Most possess a unique form of organelle structure that comprises a type of (non-photosynthetic) plastid called an apicoplast—with an apical complex membrane. The organelle's apical shape is an adaptation that the apicomplexan applies in penetrating a host cell.
A chloroplast is a type of membrane-bound organelle known as a plastid that conducts photosynthesis mostly in plant and algal cells. The photosynthetic pigment chlorophyll captures the energy from sunlight, converts it, and stores it in the energy-storage molecules ATP and NADPH while freeing oxygen from water in the cells. The ATP and NADPH is then used to make organic molecules from carbon dioxide in a process known as the Calvin cycle. Chloroplasts carry out a number of other functions, including fatty acid synthesis, amino acid synthesis, and the immune response in plants. The number of chloroplasts per cell varies from one, in unicellular algae, up to 100 in plants like Arabidopsis and wheat.
The dinoflagellates are a monophyletic group of single-celled eukaryotes constituting the phylum Dinoflagellata and are usually considered protists. Dinoflagellates are mostly marine plankton, but they also are common in freshwater habitats. Their populations vary with sea surface temperature, salinity, and depth. Many dinoflagellates are photosynthetic, but a large fraction of these are in fact mixotrophic, combining photosynthesis with ingestion of prey.
The alveolates are a group of protists, considered a major clade and superphylum within Eukarya. They are currently grouped with the stramenopiles and Rhizaria among the protists with tubulocristate mitochondria into the SAR supergroup.
A plastid, pl.plastids, is a membrane-bound organelle found in the cells of plants, algae, and some other eukaryotic organisms;. They are considered to be intracellular endosymbiotic cyanobacteria.
Chromista is a proposed but polyphyletic biological kingdom, refined from the Chromalveolata, consisting of single-celled and multicellular eukaryotic species that share similar features in their photosynthetic organelles (plastids). It includes all eukaryotes whose plastids contain chlorophyll c and are surrounded by four membranes. If the ancestor already possessed chloroplasts derived by endosymbiosis from red algae, all non-photosynthetic Chromista have secondarily lost the ability to photosynthesise. Its members might have arisen independently as separate evolutionary groups from the last eukaryotic common ancestor.
The Syndiniales are an order of early branching dinoflagellates, found as parasites of crustaceans, fish, algae, cnidarians, and protists. The trophic form is often multinucleate, and ultimately divides to form motile spores, which have two flagella in typical dinoflagellate arrangement. They lack a theca and chloroplasts, and unlike all other orders, the nucleus is never a dinokaryon. A well-studied example is Amoebophrya, which is a parasite of other dinoflagellates and may play a part in ending red tides. Several MALV groups have been assigned to Syndiniales; recent studies, however, show paraphyly of MALVs suggesting that only those groups that branch as sister to dinokaryotes belong to Syndiniales.
Chromalveolata was a eukaryote supergroup present in a major classification of 2005, then regarded as one of the six major groups within the eukaryotes. It was a refinement of the kingdom Chromista, first proposed by Thomas Cavalier-Smith in 1981. Chromalveolata was proposed to represent the organisms descended from a single secondary endosymbiosis involving a red alga and a bikont. The plastids in these organisms are those that contain chlorophyll c.
The Archaeplastida are a major group of eukaryotes, comprising the photoautotrophic red algae (Rhodophyta), green algae, land plants, and the minor group glaucophytes. It also includes the non-photosynthetic lineage Rhodelphidia, a predatorial (eukaryotrophic) flagellate that is sister to the Rhodophyta, and probably the microscopic picozoans. The Archaeplastida have chloroplasts that are surrounded by two membranes, suggesting that they were acquired directly through a single endosymbiosis event by phagocytosis of a cyanobacterium. All other groups which have chloroplasts, besides the amoeboid genus Paulinella, have chloroplasts surrounded by three or four membranes, suggesting they were acquired secondarily from red or green algae. Unlike red and green algae, glaucophytes have never been involved in secondary endosymbiosis events.
An apicoplast is a derived non-photosynthetic plastid found in most Apicomplexa, including Toxoplasma gondii, and Plasmodium falciparum and other Plasmodium spp., but not in others such as Cryptosporidium. It originated from algae through secondary endosymbiosis; there is debate as to whether this was a green or red alga. The apicoplast is surrounded by four membranes within the outermost part of the endomembrane system. The apicoplast hosts important metabolic pathways like fatty acid synthesis, isoprenoid precursor synthesis and parts of the heme biosynthetic pathway.
Myzozoa is a grouping of specific phyla within Alveolata, that either feed through myzocytosis, or were ancestrally capable of feeding through myzocytosis.
Amphidinium is a genus of dinoflagellates. The type for the genus is Amphidinium operculatum Claparède & Lachmann. The genus includes the species Amphidinium carterae which is used as a model organism.
The N-end rule is a rule that governs the rate of protein degradation through recognition of the N-terminal residue of proteins. The rule states that the N-terminal amino acid of a protein determines its half-life. The rule applies to both eukaryotic and prokaryotic organisms, but with different strength, rules, and outcome. In eukaryotic cells, these N-terminal residues are recognized and targeted by ubiquitin ligases, mediating ubiquitination thereby marking the protein for degradation. The rule was initially discovered by Alexander Varshavsky and co-workers in 1986. However, only rough estimations of protein half-life can be deduced from this 'rule', as N-terminal amino acid modification can lead to variability and anomalies, whilst amino acid impact can also change from organism to organism. Other degradation signals, known as degrons, can also be found in sequence.
Goussia is a taxonomic genus, first described in 1896 by Labbé, containing parasitic protists which largely target fish and amphibians as their hosts. Members of this genus are homoxenous and often reside in the gastrointestinal tract of the host, however others may be found in organs such as the gallbladder or liver. The genera Goussia, as current phylogenies indicate, is part of the class Conoidasida, which is a subset of the parasitic phylum Apicomplexa; features of this phylum, such as a distinct apical complex containing specialized secretory organelles, an apical polar ring, and a conoid are all present within Goussia, and assist in the mechanical invasion of host tissue. The name Goussia is derived from the French word gousse, meaning pod. This name is based on the bi-valve sporocyst morphology which some Goussians display. Of the original 8 classified Goussians, 6 fit the “pod” morphology. As of this writing, the genera consists of 59 individual species.
Vitrella brassicaformis (CCMP3155) is a unicellular alga belonging to the eukaryotic supergroup Alveolata. V. brassicaformis and its closest known relative, Chromera velia, are the only two currently described members of the phylum Chromerida, which in turn constitutes part of the taxonomically unranked group Colpodellida. Chromerida is phylogenetically closely related to the phylum Apicomplexa, which includes Plasmodium, the agent of malaria. Notably, both V. brassicaformis and C. velia are photosynthetic, each containing a complex secondary plastid. This characteristic defined the discovery of these so-called 'chromerids,' as their photosynthetic capacity positioned them to shed light upon the evolution of Apicomplexa's non-photosynthetic parasitism. Both genera lack chlorophyll b or c; these absences link the two taxonomically, as algae bearing only chlorophyll a are rare amid the biodiversity of life. Despite their similarities, V. brassicaformis differs significantly from C. velia in morphology, lifecycle, and accessory photosynthetic pigmentation. V. brassicaformis has a green color, with a complex lifecycle involving multiple pathways and a range of sizes and morphologies, while Chromera has a brown color and cycles through a simpler process from generation to generation. The color differences are due to differences in accessory pigments.
Plesiastrea versipora is an encrusting coral found in the Indian and Pacific Oceans. It is of interest because of its ability to thrive in both tropical and temperate environments, and to grow massive.
Floridean starch is a type of a storage glucan found in glaucophytes and in red algae, in which it is usually the primary sink for fixed carbon from photosynthesis. It is found in grains or granules in the cell's cytoplasm and is composed of an α-linked glucose polymer with a degree of branching intermediate between amylopectin and glycogen, though more similar to the former. The polymers that make up floridean starch are sometimes referred to as "semi-amylopectin".
Durinskia is a genus of dinoflagellates that can be found in freshwater and marine environments. This genus was created to accommodate its type species, Durinskia baltica, after major classification discrepancies were found. While Durinskia species appear to be typical dinoflagellates that are armored with cellulose plates called theca, the presence of a pennate diatom-derived tertiary endosymbiont is their most defining characteristic. This genus is significant to the study of endosymbiotic events and organelle integration since structures and organelle genomes in the tertiary plastids are not reduced. Like some dinoflagellates, species in Durinskia may cause blooms.
Rhodelphis is a single-celled archaeplastid that lives in aquatic environments and is the sister group to red algae and possibly Picozoa. While red algae have no flagellated stages and are generally photoautotrophic, Rhodelphis is a flagellated predator containing a non-photosynthetic plastid. This group is important to the understanding of plastid evolution because they provide insight into the morphology and biochemistry of early archaeplastids. Rhodelphis contains a remnant plastid that is not capable of photosynthesis, but may play a role in biochemical pathways in the cell like heme synthesis and iron-sulfur clustering. The plastid does not have a genome, but genes are targeted to it from the nucleus. Rhodelphis is ovoid with a tapered anterior end bearing two perpendicularly-oriented flagella.
Chrompodellids are a clade of single-celled protists belonging to the Alveolata supergroup. It comprises two different polyphyletic groups of flagellates: the colpodellids, phagotrophic predators, and the chromerids, photosynthetic algae that live as symbionts of corals. These groups were independently discovered and described, but molecular phylogenetic analyses demonstrated that they are intermingled in a clade that is the closest relative to Apicomplexa, and they became collectively known as chrompodellids. Due to the history of their research, they are variously known in biological classification as Chromerida or Colpodellida (ICZN)/Colpodellales (ICN).
References
1 2 3 4 5 6 Oborník, Miroslav; Vancová, Marie; Lai, De-Hua; Janouškovec, Jan; Keeling, Patrick J.; Lukeš, Julius (2011). "Morphology and Ultrastructure of Multiple Life Cycle Stages of the Photosynthetic Relative of Apicomplexa, Chromera velia". Protist. 162 (1): 115–30. doi:10.1016/j.protis.2010.02.004. PMID20643580.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 Moore, Robert B.; Oborník, Miroslav; Janouškovec, Jan; Chrudimský, Tomáš; Vancová, Marie; Green, David H.; Wright, Simon W.; Davies, Noel W.; Bolch, Christopher J. S.; Heimann, Kirsten; Šlapeta, Jan; Hoegh-Guldberg, Ove; Logsdon, John M.; Carter, Dee A. (2008). "A photosynthetic alveolate closely related to apicomplexan parasites". Nature. 451 (7181): 959–63. Bibcode:2008Natur.451..959M. doi:10.1038/nature06635. PMID18288187. S2CID28005870.
↑ Oborník, Miroslav; Janouškovec, Jan; Chrudimský, Tomáš; Lukeš, Julius (2009). "Evolution of the apicoplast and its hosts: From heterotrophy to autotrophy and back again". International Journal for Parasitology. 39 (1): 1–12. doi:10.1016/j.ijpara.2008.07.010. PMID18822291.
1 2 Oborník, M; Modrý, D; Lukeš, M; Cernotíková-Stříbrná, E; Cihlář, J; Tesařová, M; Kotabová, E; Vancová, M; Prášil, O; Lukeš, J (2012). "Morphology, ultrastructure and life cycle of Vitrella brassicaformis n. sp., n. gen., a novel chromerid from the Great Barrier Reef". Protist. 163 (2): 306–323. doi:10.1016/j.protis.2011.09.001. PMID22055836.
↑ Weatherby, Kate; Murray, Shauna; Carter, Dee; Šlapeta, Jan (2011). "Surface and Flagella Morphology of the Motile Form of Chromera velia Revealed by Field-Emission Scanning Electron Microscopy". Protist. 162 (1): 142–53. doi:10.1016/j.protis.2010.02.003. PMID20643581.
1 2 3 Portman, Neil; Šlapeta, Jan (2014). "The flagellar contribution to the apical complex: a new tool for the eukaryotic Swiss Army knife?". Trends in Parasitology. 30 (2): 58–64. doi:10.1016/j.pt.2013.12.006. PMID24411691.
↑ Oborník, Miroslav; Lukeš, Julius (2015). "The Organellar Genomes ofChromeraandVitrella, the Phototrophic Relatives of Apicomplexan Parasites". Annual Review of Microbiology. 69: 129–44. doi:10.1146/annurev-micro-091014-104449. PMID26092225.
↑ Yubuki, Naoji; Čepička, Ivan; Leander, Brian S. (2016). "Evolution of the microtubular cytoskeleton (flagellar apparatus) in parasitic protists". Molecular and Biochemical Parasitology. 209 (1–2): 26–34. doi:10.1016/j.molbiopara.2016.02.002. PMID26868980.
1 2 Cumbo, Vivian R.; Baird, Andrew H.; Moore, Robert B.; Negri, Andrew P.; Neilan, Brett A.; Salih, Anya; van Oppen, Madeleine J.H.; Wang, Yan; Marquis, Christopher P. (2013). "Chromera velia is Endosymbiotic in Larvae of the Reef Corals Acropora digitifera and A. tenuis". Protist. 164 (2): 237–44. doi:10.1016/j.protis.2012.08.003. PMID23063731.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.