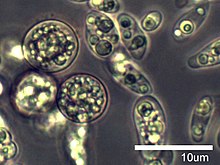
The opisthokonts are a broad group of eukaryotes, including both the animal and fungus kingdoms. The opisthokonts, previously called the "Fungi/Metazoa group", are generally recognized as a clade. Opisthokonts together with Apusomonadida and Breviata comprise the larger clade Obazoa.

The Ichthyosporea are a small group of Opisthokonta in Eukaryota, mostly parasites of fish and other animals.
Glomeromycota are one of eight currently recognized divisions within the kingdom Fungi, with approximately 230 described species. Members of the Glomeromycota form arbuscular mycorrhizas (AMs) with the thalli of bryophytes and the roots of vascular land plants. Not all species have been shown to form AMs, and one, Geosiphon pyriformis, is known not to do so. Instead, it forms an endocytobiotic association with Nostoc cyanobacteria. The majority of evidence shows that the Glomeromycota are dependent on land plants for carbon and energy, but there is recent circumstantial evidence that some species may be able to lead an independent existence. The arbuscular mycorrhizal species are terrestrial and widely distributed in soils worldwide where they form symbioses with the roots of the majority of plant species (>80%). They can also be found in wetlands, including salt-marshes, and associated with epiphytic plants.

The Laboulbeniomycetes are a unique group of fungi that are obligatorily associated with arthropods, either as external parasites or for dispersal (Pyxidiophorales).

The Laboulbeniales is an order of Fungi within the class Laboulbeniomycetes. They are also known by the colloquial name beetle hangers or labouls. The order includes around 2,325 species of obligate insect ectoparasites that produce cellular thalli from two-celled ascospores. Of the described Laboulbeniales, Weir and Hammond 1997 find 80% to be from Coleoptera and the next largest group to be the 10% from Diptera. Recently, the genus Herpomyces, traditionally considered a basal member of Laboulbeniales, was transferred to the order Herpomycetales based on molecular phylogenetic data. Laboulbeniales typically do not kill their hosts, although they may impair host fitness if the parasite density is high.

Blastocladiomycota is one of the currently recognized phyla within the kingdom Fungi. Blastocladiomycota was originally the order Blastocladiales within the phylum Chytridiomycota until molecular and zoospore ultrastructural characters were used to demonstrate it was not monophyletic with Chytridiomycota. The order was first erected by Petersen for a single genus, Blastocladia, which was originally considered a member of the oomycetes. Accordingly, members of Blastocladiomycota are often referred to colloquially as "chytrids." However, some feel "chytrid" should refer only to members of Chytridiomycota. Thus, members of Blastocladiomycota are commonly called "blastoclads" by mycologists. Alternatively, members of Blastocladiomycota, Chytridiomycota, and Neocallimastigomycota lumped together as the zoosporic true fungi. Blastocladiomycota contains 5 families and approximately 12 genera. This early diverging branch of kingdom Fungi is the first to exhibit alternation of generations. As well, two (once) popular model organisms—Allomyces macrogynus and Blastocladiella emersonii—belong to this phylum.

The Zoopagomycotina are a subdivision of the fungal division Zygomycota sensu lato. It contains 5 families and 20 genera. Relationships among and within subphyla of Zygomycota are poorly understood, and their monophyly remains in question, so they are sometimes referred to by the informal name zygomycetes.
The Harpellales are an order of fungi classified in the subdivision Kickxellomycotina. Thalli are either unbranched or branched, producing basipetal series of trichospores. Zygospores are biconical. Species in the order are found attached to the gut lining of aquatic larvae of Insecta or (rarely) Isopoda. Harpellales are divided into two families, the Harpellaceae and the Legeriomycetaceae. According to the Dictionary of the Fungi, the order contains 38 genera and 200 species. The order was formally described in 1978 Mycotaxon publication. Harpellales has served as a model to study and understand the evolution, growth, and biodiversity of other such fungi found in the gut as species are plentiful around the world.

Corallochytrium belongs to the class of Corallochytrea within Teretosporea and is a sister group to Ichthyosporea. Corallochytrium limacisporum is the only species of Corallochytrium known so far. It was first discovered and named in the Arabian Sea’s coral lagoons by Kaghu-Kumar in 1987. It was first thought to be a member of the fungi-like thraustochytrids, however, this was later disproven due to Corallochytriums lack of cilia and sagenogenetosome. Little research has been done on the life cycle or morphology. Most research concerning this genus has been done to uncover the evolution of animals and fungi, as Corallochytrium possess both animal and fungal enzymatic trademarks.

Ichthyophonida is an order of parasitic eukaryotes.

Holozoa is a clade of organisms that includes animals and their closest single-celled relatives, but excludes fungi and all other organisms. Together they amount to more than 1.5 million species of purely heterotrophic organisms, including around 300 unicellular species. It consists of various subgroups, namely Metazoa and the protists Choanoflagellata, Filasterea, Pluriformea and Ichthyosporea. Along with fungi and some other groups, Holozoa is part of the Opisthokonta, a supergroup of eukaryotes. Choanofila was previously used as the name for a group similar in composition to Holozoa, but its usage is discouraged now because it excludes animals and is therefore paraphyletic.

Amoebidiidae is a family of single-celled eukaryotes, previously thought to be zygomycete fungi belonging to the class Trichomycetes, but molecular phylogenetic analyses place the family with the opisthokont group Mesomycetozoea. The family was originally called Amoebidiaceae, and considered the sole family of the fungal order Amoebidiales that included two genera: Amoebidium and Paramoebidium. However, Amoebidiidae is now monogeneric as it was recently emended to include only Amoebidium. Species of Amoebidium are considered obligate symbionts of freshwater-dwelling arthropod hosts such as midge larvae and water fleas (Daphnia). However, because Amoebidium species attach to the exoskeleton (exterior) of the host and grow in axenic culture, at least some species may be facultative symbionts.
Smittium is a genus of fungi in the order Harpellales. It is the largest genus in the order. As of 2013, there were 81 described species. Many of these have been formally described only recently; in 1998 there were just 46. Several have been transferred to Smittium from other genera, such as Orphella, Rubetella, Genistella, and Typhella. In general, the genus has a cosmopolitan distribution, but some species are limited to small regions.
Howard C. Whisler (1931–2007) was an American mycologist. Born in Oakland, California, he attended Berkeley schools and then Palo Alto High School. Howard worked on his undergraduate degree at Oregon State College for two years and then went to the University of California, Berkeley, where he completed a Bachelor of Science degree in plant pathology in 1954. He joined the United States Air Force from 1954 to 1956 stationed in Italy. He returned to University of California, Berkeley after his military life and had finished his doctoral degree with Ralph Emerson in 1960. From 1960 to 1961 he held a post doctoral NATO-NSF Fellowship in France, at the Université de Montpellier. Howard was appointed assistant professor of Botany at McGill University in 1961. He was appointed to the faculty at the University of Washington on March 15, 1963 and worked until he died on September 16, 2007, at the age of 76.

Trichomycetes refers to a group of fungi in the division Zygomycota that grow in the guts of arthropods living in aquatic habitats. The name is obsolete, having not been validly published. Species formerly placed in the Trichomycetes are now placed in the orders Harpellales and Asellariales, both in the suborder subdivision Kickxellomycotina, while Amoebidiales and Eccrinales are included in Opisthokonta.
Robert W. Lichtwardt was a Brazilian-born American mycologist specializing in the study of arthropod-associated, gut-dwelling fungi (trichomycetes). He is known for his online monograph and interactive keys to trichomycete taxa.

Paramoebidium is a genus of unicellular, symbiotic eukaryotes that inhabit the digestive tract of immature freshwater arthropod hosts. Paramoebidium is classified in the opisthokont class Mesomycetozoea, and is the sole genus in the family Paramoebidiidae. Prior to 2005, Paramoebidium species were tentatively placed with the fungal group Trichomycetes due to their habitation of arthropod guts, host overlap between various Paramoebidium and fungal trichomycete taxa, and similar vegetative growth form.
Paramoebidiidae is a family of single-celled eukaryotes, previously thought to be zygomycete fungi belonging to the class Trichomycetes, but molecular phylogenetic analyses place the family with the opisthokont group Mesomycetozoea. The family was originally called Amoebidiaceae, and considered the sole family of the fungal order Amoebidiales that included two genera, Amoebidium and Paramoebidium. However, Paramoebidium is now the sole genus of the family Paramoebidiidae and Amoebidiidae is likewise monogeneric as it was recently emended to include only Amoebidium. Species of Paramoebidium are obligate symbionts of immature freshwater-dwelling arthropod hosts such as mayfly and stonefly nymphs and black fly larvae. Paramoebidium species attach to the digestive tract lining of their host via a secreted holdfast.
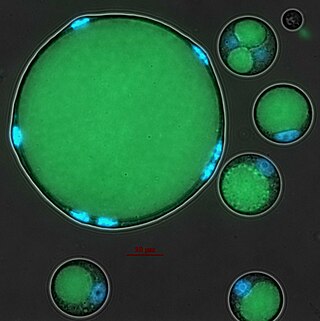
Abeoforma whisleri is a single-celled eukaryote that belongs to the Ichthyosporea clade, a group of protists closely related to animals.

Pirum gemmata is a unicellular eukaryote that belongs to the Ichthyosporea clade, a group of protists closely related to animals. P. gemmata was isolated from the gut contents of a marine invertebrate, specifically the detritivorous peanut worm Phascolosoma agassizii.