
The cell is the basic structural and functional unit of all forms of life. Every cell consists of cytoplasm enclosed within a membrane; many cells contain organelles, each with a specific function. The term comes from the Latin word cellula meaning 'small room'. Most cells are only visible under a microscope. Cells emerged on Earth about 4 billion years ago. All cells are capable of replication, protein synthesis, and motility.

A flagellum is a hairlike appendage that protrudes from certain plant and animal sperm cells, from fungal spores (zoospores), and from a wide range of microorganisms to provide motility. Many protists with flagella are known as flagellates.
The evolution of flagella is of great interest to biologists because the three known varieties of flagella – each represent a sophisticated cellular structure that requires the interaction of many different systems.

Excavata is an extensive and diverse but paraphyletic group of unicellular Eukaryota. The group was first suggested by Simpson and Patterson in 1999 and the name latinized and assigned a rank by Thomas Cavalier-Smith in 2002. It contains a variety of free-living and symbiotic protists, and includes some important parasites of humans such as Giardia and Trichomonas. Excavates were formerly considered to be included in the now obsolete Protista kingdom. They were distinguished from other lineages based on electron-microscopic information about how the cells are arranged. They are considered to be a basal flagellate lineage.

The parabasalids are a group of flagellated protists within the supergroup Excavata. Most of these eukaryotic organisms form a symbiotic relationship in animals. These include a variety of forms found in the intestines of termites and cockroaches, many of which have symbiotic bacteria that help them digest cellulose in woody plants. Other species within this supergroup are known parasites, and include human pathogens.
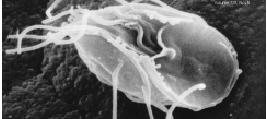
The metamonads are a large group of flagellate amitochondriate microscopic eukaryotes. Their composition is not entirely settled, but they include the retortamonads, diplomonads, and possibly the parabasalids and oxymonads as well. These four groups are all anaerobic, occurring mostly as symbiotes or parasites of animals, as is the case with Giardia lamblia which causes diarrhea in mammals.

In molecular biology, an axoneme, also called an axial filament, is the microtubule-based cytoskeletal structure that forms the core of a cilium or flagellum. Cilia and flagella are found on many cells, organisms, and microorganisms, to provide motility. The axoneme serves as the "skeleton" of these organelles, both giving support to the structure and, in some cases, the ability to bend. Though distinctions of function and length may be made between cilia and flagella, the internal structure of the axoneme is common to both.

The retortamonads are a small group of flagellates, most commonly found in the intestines of animals as commensals, although a free-living species called the Chilomastix cuspidata exists. They are grouped under the taxon, Archezoa. They are usually around 5-20 μm in length, and all of their small subunit ribosomal RNA gene sequences are very similar to each other. There are two genera: Retortamonas with two flagella, and Chilomastix with four. In both cases there are four basal bodies anterior to a prominent feeding groove, and one flagellum is directed back through the cell, emerging from the groove.
The radial spoke is a multi-unit protein structure found in the axonemes of eukaryotic cilia and flagella. Although experiments have determined the importance of the radial spoke in the proper function of these organelles, its structure and mode of action remain poorly understood.

Malawimonadidae is a family of unicellular eukaryotes of outsize importance in understanding eukaryote phylogeny.
Trimastix is a genus of excavate protists, the sole occupant of the order Trimastigida. Trimastix are bacterivorous, free living and anaerobic. It was first observed in 1881 by William Kent. There are few known species, and the genus's role in the ecosystem is largely unknown. However, it is known that they generally live in marine environments within the tissues of decaying organisms to maintain an anoxic environment. Much interest in this group is related to its close association with other members of Preaxostyla. These organisms do not have classical mitochondria, and as such, much of the research involving these microbes is aimed at investigating the evolution of mitochondria.
Malawimonas is genus of unicellular, heterotrophic flagellates with uncertain phylogenetic affinities. They have variably being assigned to Excavata and Loukozoa. Recent studies suggest they may be closely related to the Podiata.
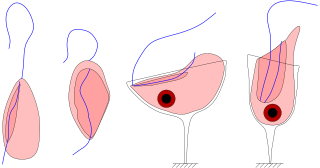
Jakobids are an order of free-living, heterotrophic, flagellar eukaryotes in the supergroup Excavata. They are small, and can be found in aerobic and anaerobic environments. The order Jakobida, believed to be monophyletic, consists of only twenty species at present, and was classified as a group in 1993. There is ongoing research into the mitochondrial genomes of jakobids, which are unusually large and bacteria-like, evidence that jakobids may be important to the evolutionary history of eukaryotes.
Jakoba is a genus in the taxon Excavata, and currently has a single described species, Jakoba libera described by Patterson in 1990, and named in honour of Dutch botanist Jakoba Ruinen.
Monocercomonoides is a genus of flagellate Excavata belonging to the order Oxymonadida. It was established by Bernard V. Travis and was first described as those with "polymastiginid flagellates having three anterior flagella and a trailing one originating at a single basal granule located in front of the anteriorly positioned nucleus, and a more or less well-defined axostyle". It is the first eukaryotic genus to be found to completely lack mitochondria, and all hallmark proteins responsible for mitochondrial function. The genus also lacks any other mitochondria related organelles (MROs) such as hydrogenosomes or mitosomes. Data suggests that the absence of mitochondria is not an ancestral feature, but rather due to secondary loss. Monocercomonoides sp. was found to obtain energy through an enzymatic action of nutrients absorbed from the environment. The genus has replaced the iron-sulfur cluster assembly pathway with a cytosolic sulfur mobilization system, likely acquired by horizontal gene transfer from a eubacterium of a common ancestor of oxymonads. These organisms are significant because they undermine assumptions that eukaryotes must have mitochondria to properly function. The genome of Monocercomonoides exilis has approximately 82 million base pairs, with 18 152 predicted protein-coding genes.
Colponema is a genus of single-celled flagellates that feed on eukaryotes in aquatic environments and soil. The genus contains 6 known species and has not been thoroughly studied. Colponema has two flagella which originate just below the anterior end of the cell. One extends forwards and the other runs through a deep groove in the surface and extends backwards. Colponema is a predator that feeds on smaller flagellates using its ventral groove. Like many other alveolates, they possess trichocysts, tubular mitochondrial cristae, and alveoli. It has been recently proposed that Colponema may be the sister group to all other alveolates. The genus could help us understand the origin of alveolates and shed light on features that are ancestral to all eukaryotes.
Stygiella /ˌstɪ.d͡ʒiˈɛ.lə/ is a genus of free-living marine flagellates belonging to the family Stygiellidae in the jakobids (excavata).

Ultrastructural identity is a concept in biology. It asserts that evolutionary lineages of eukaryotes in general and protists in particular can be distinguished by complements and arrangements of cellular organelles. These ultrastructural components can be visualized by electron microscopy.

Colponemids are free-living alveolates, unicellular flagellates related to dinoflagellates, apicomplexans and ciliates. They are predators of other small eukaryotes, found in freshwater, marine and soil environments. They do not form a solid clade, but a sparse group of deep-branching alveolate lineages.
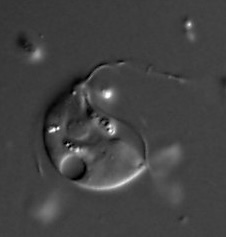
Paratrimastix pyriformis is a species of free-living anaerobic freshwater bacteriovorous flagellated protists formerly known as Trimastix pyriformis and Tetramitus pyriformis.