
Euglenozoa are a large group of flagellate Discoba. They include a variety of common free-living species, as well as a few important parasites, some of which infect humans. Euglenozoa are represented by four major groups, i.e., Kinetoplastea, Diplonemea, Euglenida, and Symbiontida. Euglenozoa are unicellular, mostly around 15–40 μm (0.00059–0.00157 in) in size, although some euglenids get up to 500 μm (0.020 in) long.

Euglena is a genus of single cell flagellate eukaryotes. It is the best known and most widely studied member of the class Euglenoidea, a diverse group containing some 54 genera and at least 200 species. Species of Euglena are found in fresh water and salt water. They are often abundant in quiet inland waters where they may bloom in numbers sufficient to color the surface of ponds and ditches green (E. viridis) or red (E. sanguinea).
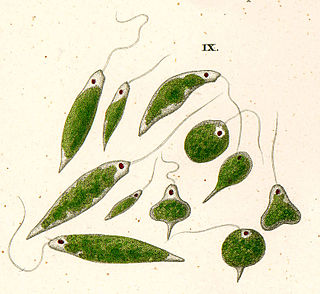
Euglenids are one of the best-known groups of flagellates, which are excavate eukaryotes of the phylum Euglenophyta and their cell structure is typical of that group. They are commonly found in freshwater, especially when it is rich in organic materials, with a few marine and endosymbiotic members. Many euglenids feed by phagocytosis, or strictly by diffusion. A monophyletic group consisting of the mixotrophic Rapaza viridis and the two groups Eutreptiales and Euglenales have chloroplasts and produce their own food through photosynthesis. This group is known to contain the carbohydrate paramylon.

Kinetoplastida is a group of flagellated protists belonging to the phylum Euglenozoa, and characterised by the presence of a distinctive organelle called the kinetoplast, a granule containing a large mass of DNA. The group includes a number of parasites responsible for serious diseases in humans and other animals, as well as various forms found in soil and aquatic environments. The organisms are commonly referred to as "kinetoplastids" or "kinetoplasts".

Excavata is an extensive and diverse but paraphyletic group of unicellular Eukaryota. The group was first suggested by Simpson and Patterson in 1999 and the name latinized and assigned a rank by Thomas Cavalier-Smith in 2002. It contains a variety of free-living and symbiotic protists, and includes some important parasites of humans such as Giardia and Trichomonas. Excavates were formerly considered to be included in the now obsolete Protista kingdom. They were distinguished from other lineages based on electron-microscopic information about how the cells are arranged. They are considered to be a basal flagellate lineage.
Trans-splicing is a special form of RNA processing where exons from two different primary RNA transcripts are joined end to end and ligated. It is usually found in eukaryotes and mediated by the spliceosome, although some bacteria and archaea also have "half-genes" for tRNAs.

Discosea is a class of Amoebozoa, consisting of naked amoebae with a flattened, discoid body shape. Members of the group do not produce tubular or subcylindrical pseudopodia, like amoebae of the class Tubulinea. When a discosean is in motion, a transparent layer called hyaloplasm forms at the leading edge of the cell. In some discoseans, short "subpseudopodia" may be extended from this hyaloplasm, but the granular contents of the cell do not flow into these, as in true pseudopodia. Discosean amoebae lack hard shells, but some, like Cochliopodium and Korotnevella secrete intricate organic scales which may cover the upper (dorsal) surface of the cell. No species have flagella or flagellated stages of life.

The Archamoebae are a group of protists originally thought to have evolved before the acquisition of mitochondria by eukaryotes. They include genera that are internal parasites or commensals of animals. A few species are human pathogens, causing diseases such as amoebic dysentery. The other genera of archamoebae live in freshwater habitats and are unusual among amoebae in possessing flagella. Most have a single nucleus and flagellum, but the giant amoeba Pelomyxa has many of each.

Bodo saltans is a free-living nonparasitic species of kinetoplastid flagellated phagotrophic protozoa that feed on bacteria. Bodo saltans cells have been reported in freshwater and marine environments.

Euglenales is an order of flagellates in the phylum Euglenozoa. The family includes the most well-known euglenoid genus, Euglena, as well as other common genera like Phacus and Lepocinclis.
Euglenaceae is a family of flagellates in the phylum Euglenozoa. The family includes the most well-known euglenoid genus, Euglena.

Malawimonadidae is a family of unicellular eukaryotes of outsize importance in understanding eukaryote phylogeny.
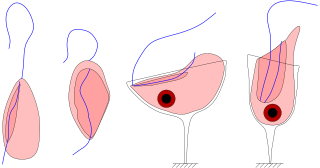
Jakobids are an order of free-living, heterotrophic, flagellar eukaryotes in the supergroup Excavata. They are small, and can be found in aerobic and anaerobic environments. The order Jakobida, believed to be monophyletic, consists of only twenty species at present, and was classified as a group in 1993. There is ongoing research into the mitochondrial genomes of jakobids, which are unusually large and bacteria-like, evidence that jakobids may be important to the evolutionary history of eukaryotes.
Cryomonadida is a group of heterotrophic Rhizaria, that belong to the Cercozoa.

Hemiselmis is a genus of cryptomonads.

Peranema is a genus of free-living phagotrophic euglenids. There are more than 20 nominal species, varying in size between 8 and 200 micrometers. Peranema cells are gliding flagellates found in freshwater lakes, ponds and ditches, and are often abundant at the bottom of stagnant pools rich in decaying organic material. Although they belong to the class Euglenoidea, and are morphologically similar to the green Euglena, Peranema have no chloroplasts, and do not conduct autotrophy. Instead, they capture live prey, such as yeast, bacteria and other flagellates, consuming them with the help of a rigid feeding apparatus called a "rod-organ." Unlike the green euglenids, they lack both an eyespot (stigma), and the paraflagellar body (photoreceptor) that is normally coupled with that organelle. However, while Peranema lack a localized photoreceptor, they do possess the light-sensitive protein rhodopsin, and respond to changes in light with a characteristic "curling behaviour."

Neobodo are diverse protists belonging to the eukaryotic supergroup Excavata. They are Kinetoplastids in the subclass Bodonidae. They are small, free-living, heterotrophic flagellates with two flagella of unequal length used to create a propulsive current for feeding. As members of Kinetoplastids, they have an evident kinetoplast There was much confusion and debate within the class Kinetoplastid and subclass Bodonidae regarding the classification of the organism, but finally the new genera Neobodo was proposed by Keith Vickerman. Although they are one of the most common flagellates found in freshwater, they are also able to tolerate saltwater Their ability to alternate between both marine and freshwater environments in many parts of the world give them a “cosmopolitan” character. Due to their relatively microscopic size ranging between 4–12 microns, they are further distinguished as heterotrophic nanoflagellates. This small size ratio limits them as bacterivores that swim around feeding on bacteria attached to surfaces or in aggregates.
Heteronema is a genus of phagotrophic, flagellated euglenoids that are most widely distributed in fresh water environments. This genus consists of two very distinguishable morphogroups that are phylogenetically closely related. These morphogroups are deciphered based on shape, locomotion and other ultrastructural traits. However, this genus does impose taxonomic problems due to the varying historical descriptions of Heteronema species and its similarity to the genus Paranema. The species H. exaratum, was the first heteronemid with a skidding motion to be sequenced, which led to the discovery that it was not closely related to H. scaphrum, contrary to what was previously assumed, but instead to a sister group of primary osmotrophs. This suggests that skidding heteronemids can also be distinguished phylogenetically, being more closely related to Anisoma, Dinema and Aphageae, than to other species within Heteronema.
Postgaardia is a proposed basal clade of flagellate Euglenozoa, following Thomas Cavalier-Smith. As of April 2023, the Interim Register of Marine and Nonmarine Genera treats the group as a subphylum. A 2021 review of Euglenozoa places Cavalier-Smith's proposed members of Postgaardia in the class Symbiontida. As Euglenozoans may be basal eukaryotes, the Postgaardia may be key to studying the evolution of Eukaryotes, including the incorporation of eukaryotic traits such as the incorporation of alphaproteobacterial mitochondrial endosymbionts.
Bihospites is a genus of symbiontid euglenozoans characterized by the presence of two species of epibiotic bacteria on the cell surface. Bihospites cells are clear, biflagellated, and uninucleated, that range between 40–120 μm long and 15–30 μm wide. Bihospites, as well as other members of the symbiontids, are found in semi-anoxic to anoxic sediments in benthic marine environments. Each cell surface is covered by both rod-shaped and spherical-shaped epibiotic bacteria that may share a commensalistic or mutualistic relationship with Bihospites host cells. Bihospites cells are highly contractile and contain several morphological synapomorphies which are present in euglenozoans, however they also contain several unique morphological traits including a unique C-shaped feeding apparatus.
