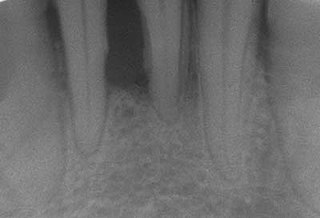
Collagen degradation observed in chronic periodontal disease results in part from the collagenase enzymes of this species. It has been shown in an in vitro study that P. gingivalis can invade humangingivalfibroblasts and can survive in the presence of antibiotics.[3]P. gingivalis invades gingival epithelialcells in high numbers, in which case both bacteria and epithelial cells survive for extended periods of time. High levels of specific antibodies can be detected in patients harboring P. gingivalis.
P. gingivalis is divided into K-serotypes based upon capsularantigenicity of the various types.[8] These serotypes have been the drivers of observations regarding bacterial cell to cell interactions to the associated serotype-dependent immune response and risk with pancreatic cancer.[9][10]
Genome
The genome of P. gingivalis was described in 2003 revealing 1,990 open reading frames (i.e. protein-coding sequences), encoded by 2,343,479 bp, with an average G+C content of 48.3%.[11] An estimated 463 genes are essential.[12]
Arg-gingipain (Rgp) and lys-gingipain (Kgp) are endopeptidase enzymes secreted by P. gingivalis. These gingipains serve many functions for the organism, contributing to its survival and virulence.[13]
Arg-gingipains have been found to play a key role in the collection of nutrients for P. gingivalis survival. Rgp degrades large peptides of the host organism to provide the bacterium with an abundant nitrogen and carbon source from human serum albumin.[14]P. gingivalis can also degrade transferrin within host cells which provides the organism with an abundant iron source needed to perform multiple cellular functions.[15]
The gingipains are also responsible for a number of necessary functions related to host invasion and colonization. Rgp gingipains are necessary for adhesion and invasion as they processed precursor proteins of long fimbriae.[15] The P. gingivalis genes encoding RgpA, Kgp, and hemagglutinin A (HagA) were strongly expressed after incubation with T. denticola. The hemagglutinin adhesion domain-containing proteins act to increase adhesive capacities of P. gingivalis with other bacterial species.[16] They are also associated with coordinating the integrity of the biofilm in the developing and maturation phase.[17] Lys- gingipains (Kgp) can bind to immobilized matrix proteins fibrinogen and fibronectin and may have a role in host colonization.[18]
Gingipains also have the ability to degrade multiple signals of the host immune response. They have the ability to cleave subclass 1 and 3 IgG antibodies[19] as well as proinflammatory cytokines such as IL-1β, IL-2, IL-6, TNF-α and IL-8 in regions of high P. gingivalis concentration,[20] impairing host immune response function. Rgp can inhibit IL-2 accumulation in T-cells, which enables it to evade the host adaptive immune response, by modulating T-cell communication and proliferation.[21]
Gingipains are key factors in tissue damage symptoms of periodontitis, which results from the degradation of matrix metalloproteins, collagen, and fibronectin.[15] Degradation of these substrates interferes with interactions between host cells and the extracellular matrix, therefore impeding wound healing and causing destruction of periodontal tissues.[15] Rgp is responsible for eliciting the host inflammatory response via the p38α MAPK transduction pathway. This response likely contributes to the inflammatory nature of periodontitis and is involved in tissue and bone destruction.[14]
Gingipains have been associated with Alzheimer's disease (AD). Gingipains were discovered from TMAs of patients exhibiting AD brain pathology. Both RgpB and Kgp were discovered from hippocampus and cerebral cortex of AD patients and were found to be associated with tau load, a marker for AD pathology and ubiquitin, which accumulates in tau tangles and amyloid beta plaques in AD brain. P. gingivalis 16S rRNA was also discovered in the cerebral cortex and csf of AD brains. Pretreatment with gingipain inhibitors protected neuron cell degradation caused by administration of gingipains in murine model.[4]
Capsular polysaccharide
The encapsulated strain of P. gingivalis is much more virulent than the nonencapsulated strain in a mouse abscess model.[22] The capsule is a capsular polysaccharide and when present down regulates cytokine production especially proinflammatory cytokines IL-1β, IL-6, IL-8, and TNF-α, indicating host evasion responses.[20][22] However, other studies have found the capsular polysaccharide to elicit host immune responses like neutrophil migration and dose and time dependent expression of cell migration chemokines like MCP-1, KC, MIP-2 and RANTES in capsular polysaccharide-challenged murine peritoneal macrophages. These conditions are likely to contribute to the inflammatory lesions observed in periodontitis.[23]
Vaccines made from capsular polysaccharide of P. gingivalis apparently impair oral bone loss in murine models. These vaccines have been able to elicit potent immune responses such as increased IgM and IgG responses that recognize whole P. gingivalis organisms.[24]
Fimbriae
Fimbriae are appendages involved in cellular attachment and greatly contribute to virulence and are found on many Gram-negative and some Gram-positive bacteria.[citation needed]
P. gingivalis virulence is heavily associated with fimbriae as they have been characterized to be key factors in adhesion, invasion, and colonization. Fimbriae are also responsible for invasion of membrane vesicles into host cells.[15] They were found to bind to cellular α5β1 integrins, which mediated adherence and impaired the homeostatic controls of host cells.[25] Fimbriae were also found to be associated with modulating β2 integrin adhesive activity for uptake by monocytes using the CD14/TLR2/PI3K signaling complex, which may contribute to intracellular evasion tactics by P. gingivalis.[26]P. gingivalis has long fimbriae, short fimbriae, and accessory components, each of which have distinct functions.[17]
Long fimbriae
Long fimbriae (FimA), also known as major fimbriae, are long, peritrichous, filamentous components.[27] They have a role in initial attachment and organization of biofilms, as they act as adhesins that mediate invasion and colonization of host cells contributing to P. gingivalis virulence.[17]
Short fimbriae
Short fimbriae (Mfa1), also known as minor fimbriae, have distinct roles from long fimbriae and are characterized to be essential for cell-cell auto aggregation and recruitment for microcolony formation.[27] Short fimbriae are involved in cell-cell adhesion with other dental commensals. It was found to coadhere and develop biofilm in conjunction with Streptococcus gordonii by interaction with SspB streptococcal surface polypeptide.[28] This interaction may be essential in the invasion of dentinal tubules by P. gingivalis.[29]
Accessory fimbriae
Fim C, D, and E accessory components associate with the main FimA protein and have a role in binding with matrix proteins and interaction with CXC-chemokine receptor 4. Loss of function experiments have confirmed that P. gingivalis mutants deficient for Fim C, D, or E have drastically attenuated virulence.[30]
Evasion of host defenses and immune responses
P. gingivalis has many ways of evading host immune responses which affects its virulence. It does this by using a combination of gingipain proteases, a capsular polysaccharide, induction of host cell proliferation, and the cleavage of chemokines responsible for neutrophil recruitment.[19][31]
Virulent P. gingivalis further modulates leukocyte recruitment by proteolysis of cytokines and chemokines that are secreted by the host cells. The arg-gingipain and lys-gingipains are responsible for this proteolysis. In a study using a mouse model, P. gingivalis was specifically found to down-regulate IL-8 induction, causing delayed neutrophil recruitment. Prevention of neutrophil recruitment may inhibit the clearance of the bacterium from the site of infection allowing for colonization.[31]P. gingivalis is able to evade opsonophagocytosis from granulocytes by using Gingipain K (Kgp) to cleave IgG 1 and 3. This further modulates immune response by impairing signaling.[19] Other studies have found that P. gingivalis can subvert the complement pathway through C5αR and C3αR, which modulates the killing capacity of leukocytes, allowing for uncontrolled bacterial growth.[31][32][33]P. gingivalis was also found to inhibit pro inflammatory and antimicrobial responses in human monocytes and mouse macrophages by fimbrial binding to CXCR4, inducing PKA signaling and inhibiting TLR-2-mediated immune response.[34]
Once in the host cells, P. gingivalis is capable of inhibiting apoptosis by modulating the JAK/Stat pathway that controls mitochondrial apoptotic pathways.[35][17] A proliferative phenotype may be beneficial to the bacterium as it provides nutrients, impairs host cell signaling, and compromises the integrity of the epithelial cell layer, allowing for invasion and colonization.[17]
Ecology
P. gingivalis plays an important role in the onset of chronic adult periodontitis.[36] Though it is found in low abundance in the oral cavity, it causes a microbial shift of the oral cavity, allowing for uncontrolled growth of the commensal microbial community. This leads to periodontitis through the disruption of the host tissue homeostasis and adaptive immune response.[37] After using laser capture microdissection plus qRT-PCR to detect P. gingivalis in human biopsies, colocalization of P. gingivalis with CD4+ T cells was observed.[38] However, the infection mechanism of T cells by P. gingivalis remains unknown.[citation needed]
P. gingivalis has been associated with increasing the virulence of other commensal bacteria in both in vivo and in vitro experiments. P. gingivalis outer membrane vesicles were found to be necessary for the invasion of epithelial cells of Tannerella forsythia.[39]P. gingivalis short fimbriae were found to be necessary for coculture biofilm formation with Streptococcus gordonii.[28] Interproximal and horizontal alveolar bone loss in mouse models are seen in coinfections involving P. gingivalis and Treponema denticola.[40] The role of P. gingivalis in periodontitis is studied using specific pathogen-free mouse models of periodontal infections. In these models, P. gingivalis inoculation causes significant bone loss, which is a significant characteristic of the disease. In contrast, germ free mice inoculated with a P. gingivalis monoinfection incur no bone loss, indicating that P. gingivalis alone cannot induce periodontitis.[31]
Pathogenesis and Cardiovascular Comorbidities
While P. gingivalis is a part of the typical microbiotic ecosystem of the oral cavity in humans, it can also become pathogenic is provided with sufficient opportunity. When this occurs, the resultant infection is known as gingivitis or periodontitis.
Periodontal disease, which an infection of the gum tissue driven primarily by P. gingivalis, has been theorized to be linked with other systemic diseases, including Alzheimer’s, atherosclerosis and other cardiovascular diseases. Though there exists no scientific consensus on the mechanism by which these disease processes are linked, evidence for the connection between periodontal disease and cardiovascular diseases including atherosclerosis has been found in both statistical study of human populations,[41] and in vivo studies using a mouse model.[42]
It is thought that periodontitis is linked to cardiovascular disease due to inflammation pathways, which the two pathologies have in common. The inflammation pathway of periodontitis is such that as the infection grows, bacteria, including P. gingivalis are targeted by neutrophils and natural killer immune cells. These cells phagocytize the bacteria, while simultaneously cytokine molecules in the area lead to a proinflammatory environment. This proinflammatory environment is also rich in intercellular signaling molecules including tumor necrosis factor-alpha, interleukins (interleukin 1, interleukin 4, interleukin 10), interferons and transforming growth factor beta. These molecules recruit more enzymes and transcription factors, which then in turn recruit more immune cells, forming a positive feedback loop which can make the immune response, and therefore the inflammation become chronic and systemic. Chronic inflammation of gum tissue can lead to a loss of this tissue and of bone tissue as well. The inflammation upregulates production of RANKL, an intercellular signaling molecule that promotes bone tissue dissolution, leading to a gradual loss of bony tissue. P. gingivalis infection is also thought to lead to oxidative stress. Both chronic systemic inflammation and oxidative stress are factors associated with the onset of cardiovascular disease, and are proposed mechanisms by which periodontal disease may, if it indeed is causationally linked to cardiovascular disease, accelerate the disease process of a cardiovascular disease.[43]
While invasive P. gingivalis is associated with various forms of cardiovascular disease, including stroke, coronary artery disease, atrial fibrillation, and heart failure, the best evidence of a direct causational link is between invasive P. gingivalis (periodontal disease) and atherosclerosis. In vivo and in vitro animal models have found that the fimbriae of P. gingivalis promote host cell entry and atherothrombotic lesion formation once the bacteria enter the bloodstream, such as through lesions in the mouth. P. gingivalis has been shown to accelerate the atherosclerosis disease pathway in mice, as well as being found in human atherosclerotic plaque lesions.[44]
Periodontal disease, also known as gum disease, is a set of inflammatory conditions affecting the tissues surrounding the teeth. In its early stage, called gingivitis, the gums become swollen and red and may bleed. It is considered the main cause of tooth loss for adults worldwide. In its more serious form, called periodontitis, the gums can pull away from the tooth, bone can be lost, and the teeth may loosen or fall out. Halitosis may also occur.
Streptococcus pneumoniae, or pneumococcus, is a Gram-positive, spherical bacteria, alpha-hemolytic member of the genus Streptococcus. They are usually found in pairs (diplococci) and do not form spores and are non motile. As a significant human pathogenic bacterium S. pneumoniae was recognized as a major cause of pneumonia in the late 19th century, and is the subject of many humoral immunity studies.
The bacterial capsule is a large structure common to many bacteria. It is a polysaccharide layer that lies outside the cell envelope, and is thus deemed part of the outer envelope of a bacterial cell. It is a well-organized layer, not easily washed off, and it can be the cause of various diseases.
Aspergillus fumigatus is a species of fungus in the genus Aspergillus, and is one of the most common Aspergillus species to cause disease in individuals with an immunodeficiency.
Dental plaque is a biofilm of microorganisms that grows on surfaces within the mouth. It is a sticky colorless deposit at first, but when it forms tartar, it is often brown or pale yellow. It is commonly found between the teeth, on the front of teeth, behind teeth, on chewing surfaces, along the gumline (supragingival), or below the gumline cervical margins (subgingival). Dental plaque is also known as microbial plaque, oral biofilm, dental biofilm, dental plaque biofilm or bacterial plaque biofilm. Bacterial plaque is one of the major causes for dental decay and gum disease.
Adhesins are cell-surface components or appendages of bacteria that facilitate adhesion or adherence to other cells or to surfaces, usually in the host they are infecting or living in. Adhesins are a type of virulence factor.
Aggregatibacter actinomycetemcomitans is a Gram-negative, facultative anaerobe, nonmotile bacterium that is often found in association with localized aggressive periodontitis, a severe infection of the periodontium. It is also suspected to be involved in chronic periodontitis. Less frequently, A. actinomycetemcomitans is associated with nonoral infections such as endocarditis. Its role in aggressive periodontitis was first discovered by Danish-born periodontist Jørgen Slots, a professor of dentistry and microbiology at the University of Southern California School of Dentistry.
Treponema denticola is a Gram-negative, obligate anaerobic, motile and highly proteolytic spirochete bacterium. It is one of four species of oral spirochetes to be reliably cultured, the others being Treponema pectinovorum, Treponema socranskii and Treponema vincentii. T. denticola dwells in a complex and diverse microbial community within the oral cavity and is highly specialized to survive in this environment. T. denticola is associated with the incidence and severity of human periodontal disease. Treponema denticola is one of three bacteria that form the Red Complex, the other two being Porphyromonas gingivalis and Tannerella forsythia. Together they form the major virulent pathogens that cause chronic periodontitis. Having elevated T. denticola levels in the mouth is considered one of the main etiological agents of periodontitis. T. denticola is related to the syphilis-causing obligate human pathogen, Treponema pallidum subsp. pallidum. It has also been isolated from women with bacterial vaginosis.
Pneumococcal infection is an infection caused by the bacterium Streptococcus pneumoniae.
Prevotella is a genus of Gram-negative bacteria.
Porphyromonas is a Gram-negative, non-spore-forming, obligately anaerobic and non-motile genus from the family Porphyromonadaceae. There were 16 different Porphyromonas species documented as of 2015, which reside in both animal and human reservoirs. It was discovered more recently that Porphyromonas also exist in the environment, albeit to a lesser extent. This genus is notably implicated in the modulation of oral cavity, respiratory tract, and gastrointestinal tract disease states. It is suggested that Porphyromonas either operate as benign bacteria pertinent to host immunity or are potential pathobionts that opportunistically provoke diseased states when homeostasis is disrupted. Despite its characterization not being fully elucidated due to sparse research, various studies report the prevalence of this genus at 58.7% in healthy states compared with 41.3% in diseased states.
Chronic periodontitis is one of the seven categories of periodontitis as defined by the American Academy of Periodontology 1999 classification system. Chronic periodontitis is a common disease of the oral cavity consisting of chronic inflammation of the periodontal tissues that is caused by the accumulation of profuse amounts of dental plaque. Periodontitis initially begins as gingivitis and can progress onto chronic and subsequent aggressive periodontitis according to the 1999 classification.
Aggressive periodontitis describes a type of periodontal disease and includes two of the seven classifications of periodontitis as defined by the 1999 classification system:
Localized aggressive periodontitis (LAP)
Generalized aggressive periodontitis (GAP)
Actinobacillus pleuropneumoniae, is a Gram-negative, facultative anaerobic, respiratory pathogen found in pigs. It was first reported in 1957, and was formally declared to be the causative agent of porcine pleuropneumonia in 1964. It was reclassified in 1983 after DNA studies showed it was more closely related to A. lignieresii.
Gingipain K is an enzyme. This enzyme catalyses the following chemical reaction
Prevotella nigrescens is a species of bacterium. Prevotella nigrescens has a gram negative gram stain. When P. nigrescens microflora colonize they trigger an over-aggressive response from the immune system and increase the incidence of many diseases and infections. One specific type of bacteria that is part of the normal oral flora but leads to disease when it infects the local tissue. This bacteria has no means of motility and has a Bacilli (rod) shape. Prevotella species are part of the human oral and vaginal flora. They play a role in the pathogenesis of periodontal disease, gingivitis, and extraoral infections such as nasopharyngeal and intra-abdominal infections also some odontogenic infections, and strains are usually carried in families, in so-called intrafamilial carriage. It is also associated with carotid atherosclerosis.
Prevotella brevis is a species of bacterium.
Prevotella bryantii, previously known as Bacteroides ruminicola subsp. brevis biovar 3, is a species of bacterium.
Plaque hypotheses are theories to explain the role of plaque bacteria in dental caries and in periodontitis. They rely heavily on the postulates of Koch and on the work of Louis Pasteur (1822–1895). Changing perceptions have altered treatment models.
P fimbriae are chaperone-usher type fimbrial appendages found on the surface of many Escherichia coli bacteria. The P fimbriae is considered to be one of the most important virulence factor in uropathogenic E. coli and plays an important role in upper urinary tract infections. P fimbriae mediate adherence to host cells, a key event in the pathogenesis of urinary tract infections.
↑ Irshad M, van der Reijden WA, Crielaard W, Laine ML (December 2012). "In vitro invasion and survival of Porphyromonas gingivalis in gingival fibroblasts; role of the capsule". Archivum Immunologiae et Therapiae Experimentalis. 60 (6): 469–76. doi:10.1007/s00005-012-0196-8. PMID22949096. S2CID14254746.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.